Modul 1 – Konsep Dasar Genetika
- Konsep Genetika
- Transkripsi dan Translasi
- Video Pembelajaran
- Quiz
- Forum
Gambar 3D Kromosom
Pengertian Genetika
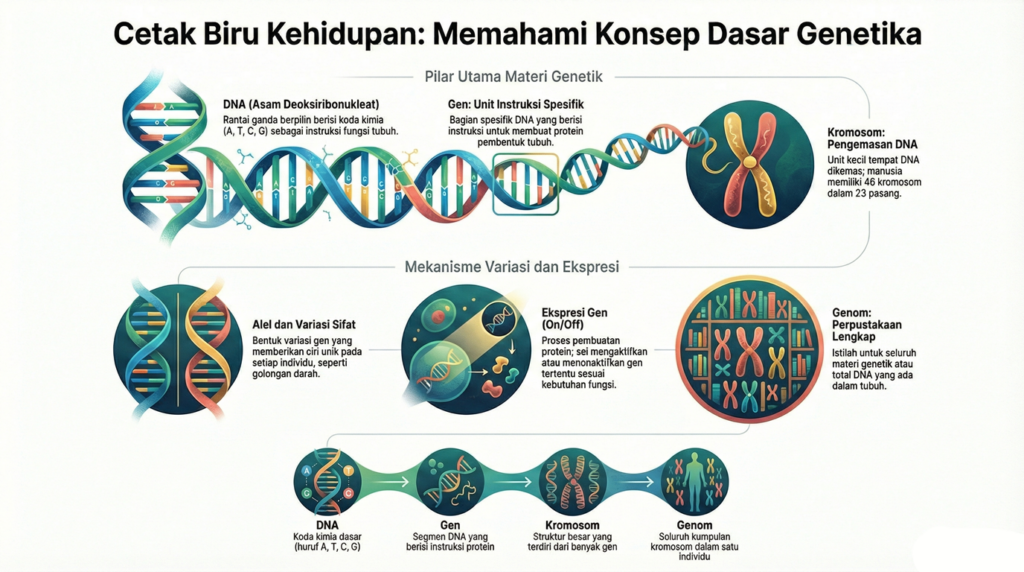
Genetika merupakan cabang ilmu biologi yang mempelajari gen, pewarisan sifat (hereditas), serta variasi karakteristik pada makhluk hidup. Genetika tradisional menekankan pada urutan atau sekuens DNA sebagai faktor utama yang menentukan sifat dan fungsi suatu organisme.
Istilah-istilah yang perlu diketahui:
DNA
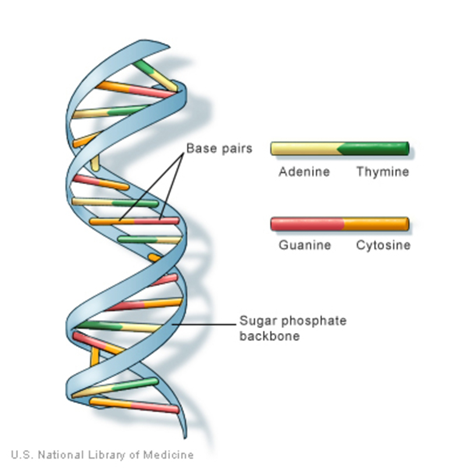
Gambar 1. DNA (Deoxyribonucleic Acid atau asam deoksiribonukleat) (Sumber: Medineplus)
DNA (singkatan dari asam deoksiribonukleat) mengandung instruksi untuk membuat tubuh Anda berfungsi. DNA terdiri dari dua untai yangsaling melilit dan terlihat seperti tangga yang berpilin (bentuk yang disebut heliks ganda). Setiap untai DNA mengandung bahan kimia yangdisebut basa nitrogen, yang membentuk kode DNA. Ada empat basa yang berbeda, T (timin), A (adenin), C (sitosin), dan G (guanin). Setiap basapada satu untai DNA berpasangan dengan basa pada untai lainnya. Basa yang berpasangan membentuk “anak tangga DNA”.
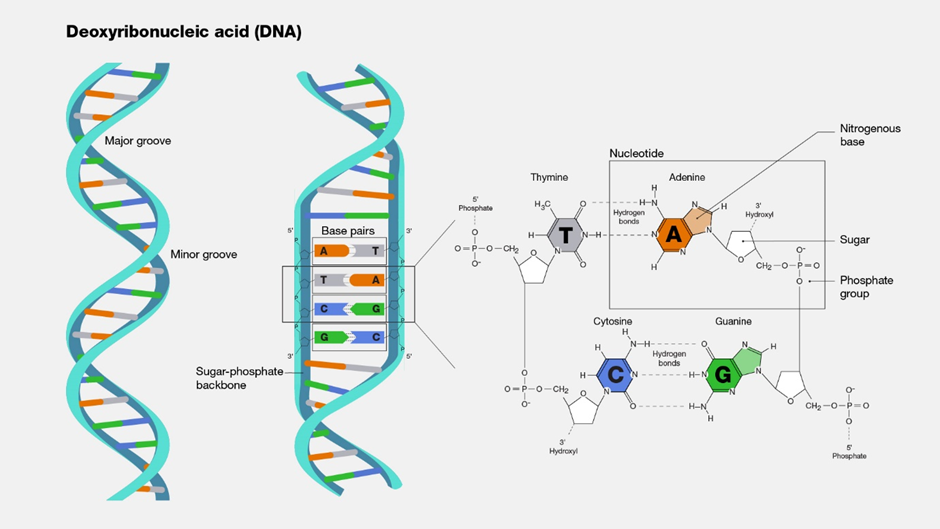
Gambar 2. Komponen DNA
(Sumber: Genetics glossary NIH)
Basa-basa tersebut tersusun dalam urutan yang berbeda di bagian-bagian untai DNA yang berbeda. DNA “dibaca” berdasarkan urutan basa,yaitu berdasarkan urutan T, C, G, dan A. Urutan basa-basa inilah yang dikenal sebagai sekuens DNA. DNA pada hampir semua makhluk hiduptersusun dari bagian-bagian yang sama. Yang berbeda adalah sekuens DNA-nya.
GENETIK
GENETIK merupakan pewarisan genetik adalah proses pewarisan DNA dari orang tua kepada anak.
GENOM
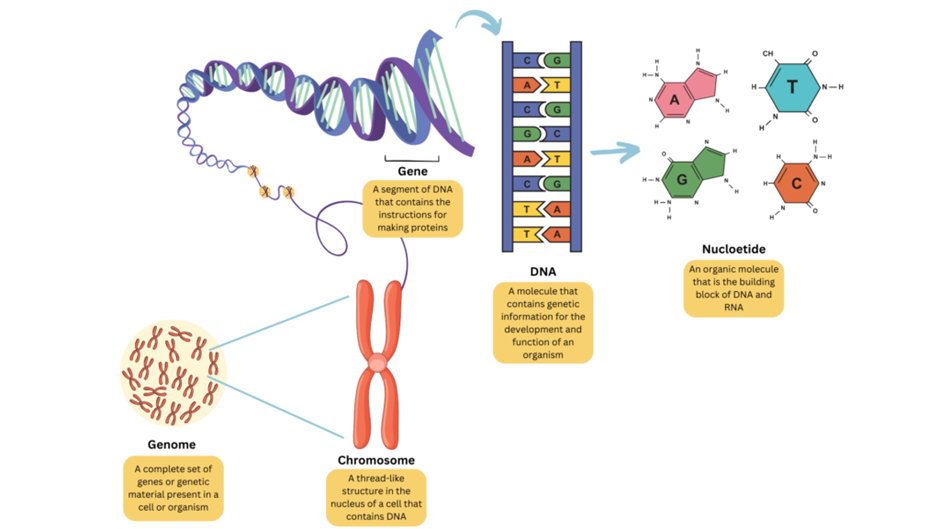
Gambar 3. Genom (Sumber: National Institute of Health)
Genom adalah seluruh DNA yang ada di dalam tubuh Anda.
Genom adalah unit penyimpanan informasi suatu organisme yang terdiri dari jutaan dan miliaran huruf DNA (A, T, C & G). Genom manusia terdiri dari sekitar 3 miliar huruf, yang mengkodekan instruksi untuk kehidupan. Informasi ini diturunkan dari satu generasi ke generasi berikutnya dan membawa sifat-sifat seperti ciri fisik dan kecenderungan penyakit. Fungsi utama genom adalah untuk menyimpan, berbagi, dan mengekspresikan informasi genetik, yang menentukan struktur dan fungsi sel. Genom manusia telah diurutkan dan dianalisis untuk memberikan wawasan tentang perkembangan manusia, fisiologi, kedokteran, dan evolusi.
Kromosom
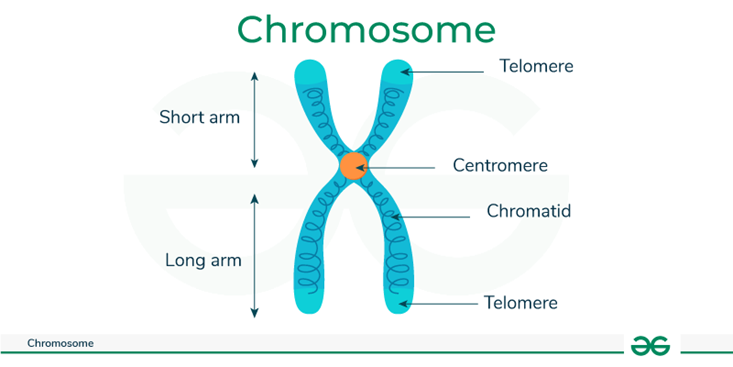
Gambar 4. Kromosom
DNA dikemas menjadi unit-unit kecil yang disebut kromosom. Sebuah kromosom mengandung satu bagian DNA yang panjang dengan banyakgen yang berbeda. Anda mewarisi kromosom dari orang tua Anda. Kromosom berpasangan. Manusia memiliki 46 kromosom, dalam 23 pasang.Anak-anak secara acak mendapatkan satu dari setiap pasangan kromosom dari ibu mereka dan satu dari setiap pasangan dari ayah mereka. Ada22 pasang kromosom bernomor, yang disebut autosom, dan kromosom yang membentuk pasangan ke-23 disebut kromosom seks. Kromosomini menentukan apakah seseorang dilahirkan sebagai laki-laki atau perempuan. Seorang perempuan memiliki dua kromosom X, dan seorang laki-laki memiliki satu kromosom X dan satu kromosom Y. Setiap anak perempuan mendapatkan satu kromosom X dari ibunya dan satu kromosom Xdari ayahnya. Setiap anak laki-laki mendapatkan satu kromosom X dari ibunya dan satu kromosom Y dari ayahnya.
Gen dan protein
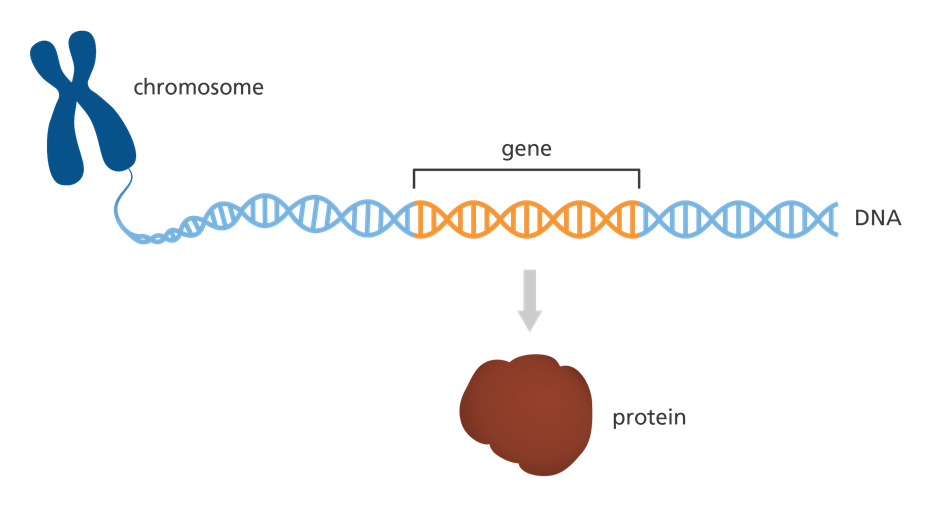
Gambar 5. Gen dan Protein
Setiap kromosom memiliki banyak gen. Gen adalah bagian spesifik dari DNA yang berisi instruksi untuk membuat protein. Protein membentuksebagian besar bagian tubuh Anda dan membuat tubuh Anda berfungsi dengan benar.
Anda memiliki dua salinan dari setiap gen. Anda mewarisi satu salinan dari ayah Anda dan satu salinan dari ibu Anda. Gen yang diwarisiseseorang dari orang tuanya dapat menentukan banyak hal. Misalnya, gen memengaruhi penampilan seseorang dan apakah orang tersebutmungkin menderita penyakit tertentu.
Alel
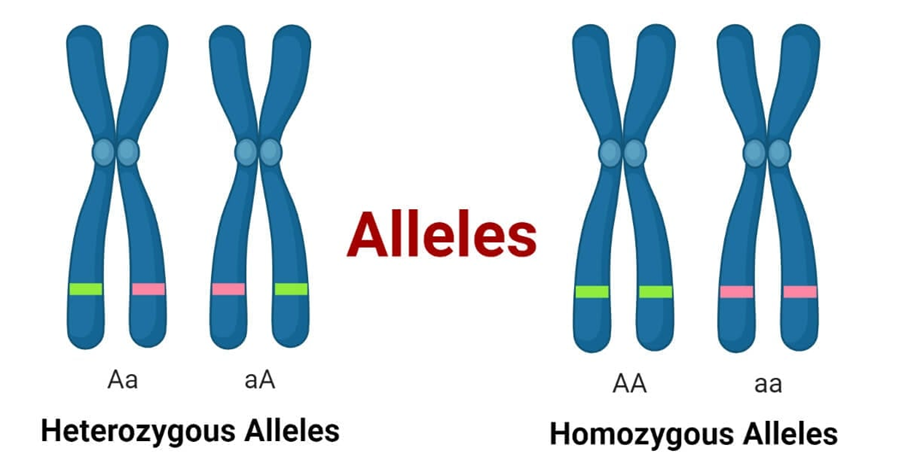
Gambar 6. Alel
Alel adalah bentuk dari gen yang sama yang mungkin memiliki sedikit perbedaan dalam urutan basa DNA-nya. Perbedaan ini berkontribusi padaciri-ciri unik setiap orang. Setiap orang memiliki dua alel untuk setiap gen, satu dari setiap orang tua. Jika alel suatu gen sama, orang tersebutdianggap homozigot untuk gen tersebut. Jika alelnya berbeda, orang tersebut dianggap heterozigot untuk gen tersebut.
Sebagian besar waktu, perbedaan antar alel tidak banyak berpengaruh pada protein yang dihasilkan. Namun, terkadang alel yang berbeda dapatmenyebabkan perbedaan sifat, seperti golongan darah. Beberapa alel dikaitkan dengan masalah kesehatan atau kelainan genetik. Pada alel-alelini, perbedaan dalam urutan basa DNA memengaruhi kemampuan tubuh untuk membuat protein tertentu.
Karena gen Anda diwariskan dari orang tua Anda, Anda dan anggota keluarga Anda memiliki banyak alel gen yang sama. Semakin dekathubungan kekerabatan Anda, semakin banyak alel gen yang Anda miliki bersama.
Sel
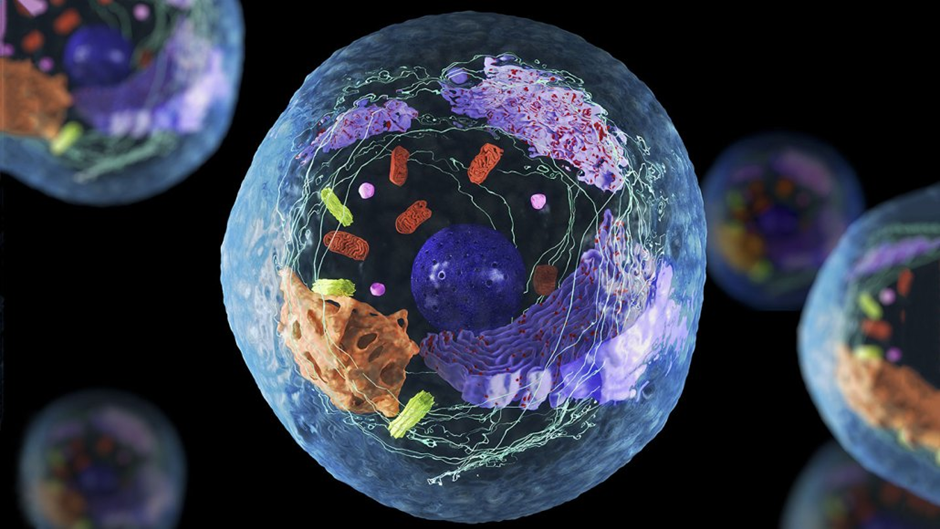
Gambar 7. Sel
Sel adalah unit dasar kehidupan. Tubuh manusia mengandung triliunan sel. Ada banyak jenis sel yang membentuk berbagai jaringan dan organdalam tubuh. Misalnya, sel kulit, sel darah, sel jantung, sel otak, dan sel ginjal hanyalah beberapa jenis sel yang menjalankan berbagai fungsivital dalam tubuh.
Struktur dasar sel adalah zat seperti jeli yang dikenal sebagai sitoplasma, yang dikelilingi oleh membran untuk menahannya agar tetap menyatu.Di dalam sitoplasma terdapat berbagai struktur khusus yang penting untuk kerja sel. Salah satu struktur ini adalah nukleus sel, yang berisi DNAyang dikemas dalam kromosom.
Copy Number Varian(CNV)

Gambar 9. Copy Number Varian (CNV)
Copy Number Varian(CNV) didefinisikan sebagai perubahan yang terjadi akibat perubahan jumlah salinan suatu gen atau gen-gen atau DNA karena translokasi, delesi, duplikasi, atau inversi. Atau pada tingkat kromosom, hal itu didefinisikan sebagai, “Variasi jumlah salinan bagian kromosom akibat penghapusan atau penggandaan.”
CNV merujuk pada ciri genom, di mana berbagai bagian DNA seseorang diulang. Meskipun ini terjadi pada semua orang,jumlah pengulangan (atau salinan) bervariasi dari satu orang ke orang lain. CNV memainkan peran penting dalam menciptakan keragamangenetik pada manusia. Namun, beberapa CNV dikaitkan dengan penyakit.
Transkripsi dan Translasi
Transkripsi dan Translasi adalah dua langkah mendasar ekspresi gen yang mengubah informasi genetik yang tersimpan dalam DNA menjadi protein fungsional Transkripsi, yang terjadi di nukleus (eukariota), mensintesis RNA dari cetakan DNA. Translasi kemudian terjadi pada ribosom di sitoplasma, menggunakan kode RNA untuk menyusun asam amino menjadi protein.
Berikut uraian tahapan masing-masing:
Traskripsi
Transkripsi terjadi di dalam nukleus. Proses ini menggunakan DNA sebagai cetakan untuk membuat molekul RNA (mRNA). Selama transkripsi, untai mRNA dibuat yang komplementer dengan untai DNA. Gambar 1 menunjukkan bagaimana proses ini terjadi.
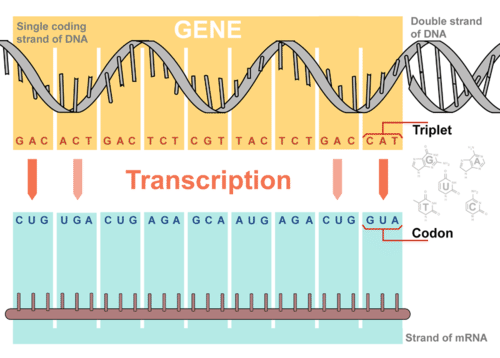
Gambar 1. Gambaran Umum Transkripsi. Transkripsi menggunakan urutan basa dalam untai DNA untuk membuat untai mRNA yang komplementer. Triplet adalah kelompok tiga basa nukleotida yang berurutan dalam DNA. Kodon adalah kelompok basa komplementer dalam mRNA.
Tahapan Transkripsi
Transkripsi berlangsung dalam tiga tahapan: inisiasi, elongasi, dan terminasi. Tahapan-tahapan tersebut diilustrasikan pada Gambar 2
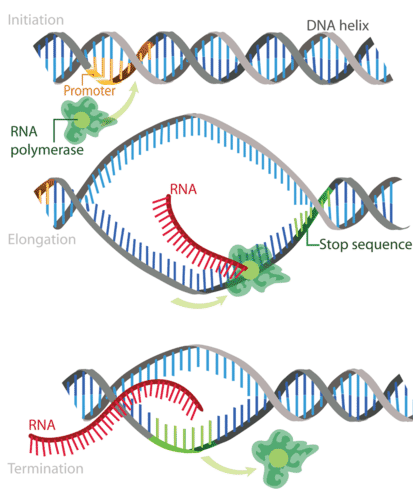
Gambar 2. Transkripsi terjadi dalam tiga langkah—inisiasi, elongasi, dan terminasi—semuanya ditunjukkan di sini.
Langkah 1: Inisiasi
Inisiasi adalah awal dari transkripsi. Ini terjadi ketika enzim RNA polimerase mengikat ke suatu wilayah gen yang disebut promotor. Hal ini memberi sinyal pada DNA untuk membuka lilitannya sehingga enzim dapat “membaca” basa-basa di salah satu untai DNA. Enzim tersebut kini siap untuk membuat untai mRNA dengan urutan basa yang komplementer
Langkah 2: Elongasi
Elongasi adalah penambahan nukleotida ke untai mRNA. RNA polimerase membaca untai DNA yang telah terurai dan membangun molekul mRNA, menggunakan pasangan basa komplementer. Ada waktu singkat selama proses ini ketika RNA yang baru terbentuk terikat pada DNA yang telah terurai. Selama proses ini, adenin (A) dalam DNA berikatan dengan urasil (U) dalam RNA
Langkah 3: Terminasi
Terminasi adalah akhir dari transkripsi, dan terjadi ketika RNA polimerase melewati sekuens berhenti (terminasi) dalam gen. Untai mRNA lengkap, dan terlepas dari DNA
RNA Polimerase
Bagian ini akan menguraikan peran spesifik RNA polimerase selama transkripsi. Lanjutkan membaca untuk mempelajari peran RNA polimerase pada setiap tahap transkripsi
Inisiasi Transkripsi
Tidak seperti polimerase prokariotik yang dapat mengikat templat DNA dengan sendirinya, eukariota membutuhkan beberapa protein lain, yang disebut faktor transkripsi, untuk terlebih dahulu mengikat wilayah promotor dan kemudian membantu merekrut polimerase yang sesuai.
Tiga Polimerase RNA Eukariotik
Ciri-ciri sintesis mRNA eukariotik jauh lebih kompleks dibandingkan prokariotik. Alih-alih satu polimerase yang terdiri dari lima subunit, eukariotik memiliki tiga polimerase yang masing-masing terdiri dari 10 subunit atau lebih. Setiap polimerase eukariotik juga membutuhkan serangkaian faktor transkripsi yang berbeda untuk membawanya ke templat DNA.
RNA polimerase I terletak di nukleolus, substruktur nuklir khusus tempat RNA ribosom (rRNA) ditranskripsikan, diproses, dan dirakit menjadi ribosom (Tabel 1). Molekul rRNA dianggap sebagai RNA struktural karena memiliki peran seluler tetapi tidak diterjemahkan menjadi protein. rRNA merupakan komponen ribosom dan penting untuk proses translasi. RNA polimerase I mensintesis semua rRNA kecuali molekul rRNA 5S. Penamaan “S” mengacu pada satuan “Svedberg”, nilai non-aditif yang mencirikan kecepatan pengendapan partikel selama sentrifugasi.

RNA polimerase II terletak di nukleus dan mensintesis semua pre-mRNA nuklear pengkode protein. Pre-mRNA eukariotik mengalami pemrosesan ekstensif setelah transkripsi tetapi sebelum translasi (Gambar 3). Untuk kejelasan, pembahasan transkripsi dan translasi pada eukariota dalam modul ini akan menggunakan istilah “mRNA” hanya untuk menggambarkan molekul matang yang telah diproses dan siap untuk ditranslasikan. RNA polimerase II bertanggung jawab untuk mentranskripsikan sebagian besar gen eukariotik.

Gambar 3. mRNA eukariotik mengandung intron yang harus dihilangkan melalui proses penyambungan (splicing). Sebuah tudung 5′ dan ekor poli-A 3′ juga ditambahkan.
RNA polimerase III juga terletak di nukleus. Polimerase ini mentranskripsikan berbagai RNA struktural yang meliputi pre-rRNA 5S, pre-RNA transfer (pre-tRNA), dan pre- RNA nuklir kecil . tRNA memiliki peran penting dalam translasi; mereka berfungsi sebagai molekul adaptor antara templat mRNA dan rantai polipeptida yang sedang tumbuh. RNA nuklir kecil memiliki berbagai fungsi, termasuk “penyambungan” pre-mRNA dan mengatur faktor transkripsi.
Seorang ilmuwan yang mengkarakterisasi gen baru dapat menentukan polimerase mana yang mentranskripsikannya dengan menguji apakah gen tersebut diekspresikan dengan adanya racun jamur tertentu, α-amanitin (Tabel 1). Menariknya, α-amanitin yang diproduksi oleh Amanita phalloides , jamur Death Cap, memengaruhi ketiga polimerase dengan sangat berbeda. RNA polimerase I sama sekali tidak sensitif terhadap α-amanitin, artinya polimerase tersebut dapat mentranskripsikan DNA secara in vitro dengan adanya racun ini. Sebaliknya, RNA polimerase II sangat sensitif terhadap α-amanitin, dan RNA polimerase III cukup sensitif. Mengetahui polimerase yang mentranskripsikan dapat memberi petunjuk kepada peneliti tentang fungsi umum gen yang sedang dipelajari. Karena RNA polimerase II mentranskripsikan sebagian besar gen, kita akan fokus pada polimerase ini dalam pembahasan selanjutnya tentang faktor transkripsi dan promotor eukariotik.
Struktur Promotor RNA Polimerase II
Promotor eukariotik jauh lebih besar dan lebih kompleks daripada promotor prokariotik, tetapi keduanya memiliki kotak TATA. Misalnya, pada gen timidina kinase tikus, kotak TATA terletak kira-kira pada posisi -30 relatif terhadap situs inisiasi (+1) (Gambar 4). Untuk gen ini, urutan kotak TATA yang tepat adalah TATAAAA, seperti yang dibaca dari arah 5′ ke 3′ pada untai non-templat. Stabilitas termal ikatan A–T rendah dan ini membantu templat DNA untuk terurai secara lokal sebagai persiapan untuk transkripsi.
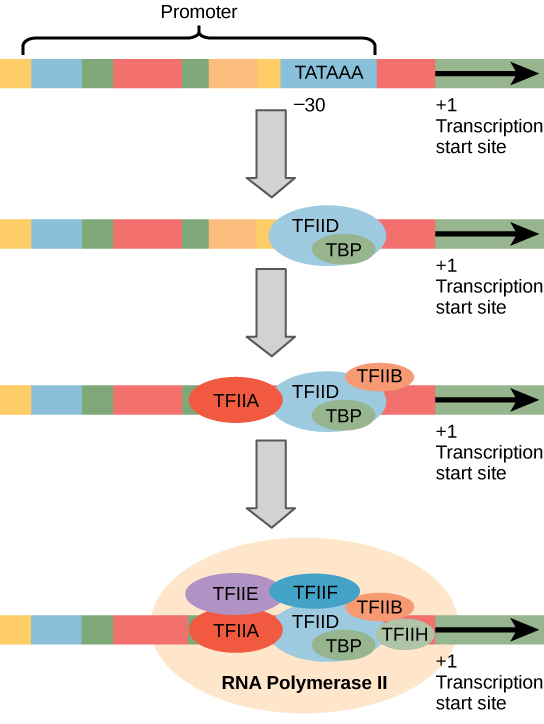
Gambar 4. Sebuah promotor umum dari gen yang ditranskripsikan oleh RNA polimerase II ditunjukkan. Faktor transkripsi mengenali promotor tersebut. RNA polimerase II kemudian mengikat dan membentuk kompleks inisiasi transkripsi.
Genom tikus mencakup satu gen dan dua pseudogen untuk timidina kinase sitoplasma. Pseudogen adalah gen yang telah kehilangan kemampuan pengkodean proteinnya atau tidak lagi diekspresikan oleh sel. Pseudogen ini disalin dari mRNA dan dimasukkan ke dalam kromosom. Misalnya, promotor timidina kinase tikus juga memiliki kotak CAAT (GGCCAATCT) yang terkonservasi pada sekitar -80. Urutan ini penting dan terlibat dalam pengikatan faktor transkripsi. Lebih jauh ke hulu dari kotak TATA, promotor eukariotik juga dapat mengandung satu atau lebih kotak kaya GC (GGCG) atau kotak oktamer (ATTTGCAT). Elemen-elemen ini mengikat faktor seluler yang meningkatkan efisiensi inisiasi transkripsi dan sering diidentifikasi dalam gen yang lebih “aktif” yang terus-menerus diekspresikan oleh sel.
Faktor Transkripsi untuk RNA Polimerase II
Kompleksitas transkripsi eukariotik tidak berakhir hanya pada polimerase dan promotor. Sejumlah besar faktor transkripsi basal, enhancer, dan silencer juga membantu mengatur frekuensi sintesis pre-mRNA dari suatu gen. Enhancer dan silencer memengaruhi efisiensi transkripsi tetapi tidak diperlukan agar transkripsi dapat berlangsung. Faktor transkripsi basal sangat penting dalam pembentukan kompleks pra-inisiasi pada templat DNA yang kemudian merekrut RNA polimerase II untuk inisiasi transkripsi.
Nama-nama faktor transkripsi basal dimulai dengan “TFII” (ini adalah faktor transkripsi untuk RNA polimerase II) dan ditentukan dengan huruf A–J. Faktor transkripsi secara sistematis menempati tempatnya pada templat DNA, dengan masing-masing faktor lebih lanjut menstabilkan kompleks pra-inisiasi dan berkontribusi pada perekrutan RNA polimerase II.
Proses membawa RNA polimerase I dan III ke templat DNA melibatkan kumpulan faktor transkripsi yang sedikit kurang kompleks, tetapi tema umumnya tetap sama. Transkripsi eukariotik adalah proses yang diatur secara ketat yang membutuhkan berbagai protein untuk berinteraksi satu sama lain dan dengan untai DNA. Meskipun proses transkripsi pada eukariota melibatkan investasi metabolik yang lebih besar daripada pada prokariota, hal ini memastikan bahwa sel mentranskripsikan secara tepat pre-mRNA yang dibutuhkannya untuk sintesis protein.
Translasi
Seperti halnya sintesis mRNA, sintesis protein dapat dibagi menjadi tiga fase: inisiasi, elongasi, dan terminasi. Proses translasi serupa pada prokariota dan eukariota. Di sini kita akan mengeksplorasi bagaimana translasi terjadi pada
E. coli , prokariota representatif, dan menjelaskan perbedaan antara translasi prokariotik dan eukariotik.
Inisiasi Translasi
Sintesis protein dimulai dengan pembentukan kompleks inisiasi. Pada E. coli , kompleks ini melibatkan ribosom 30S kecil, templat mRNA, tiga faktor inisiasi (IF; IF-1, IF-2, dan IF-3), dan tRNA inisiator khusus , yang disebut. tRNA inisiator berinteraksi dengan kodon awal AUG (atau jarang, GUG), terhubung ke metionin terformilasi yang disebut fMet, dan juga dapat mengikat IF-2. Metionin terformilasi dimasukkan oleh di awal setiap rantai polipeptida yang disintesis oleh E. coli , tetapi biasanya dipotong setelah translasi selesai. Ketika AUG sekerangkaian ditemukan selama pemanjangan translasi, metionin yang tidak terformilasi dimasukkan oleh Met-tRNA Met biasa
Dalam mRNA
E. coli , urutan di hulu kodon AUG pertama, yang disebut
urutan Shine-Dalgarno (AGGAGG), berinteraksi dengan molekul rRNA yang membentuk ribosom. Interaksi ini menambatkan subunit ribosom 30S pada lokasi yang tepat pada templat mRNA. Guanosin trifosfat (GTP), yang merupakan nukleotida purin trifosfat, bertindak sebagai sumber energi selama translasi—baik pada awal elongasi maupun selama translokasi ribosom.
Pada eukariota, kompleks inisiasi serupa terbentuk, yang terdiri dari mRNA, subunit ribosom kecil 40S, IF, dan nukleosida trifosfat (GTP dan ATP). tRNA inisiator bermuatan, yang disebut Met-tRNAi, tidak mengikat fMet pada eukariota, tetapi berbeda dari Met-tRNA lainnya karena dapat mengikat IF.
Alih-alih mengendapkan pada sekuens Shine-Dalgarno, kompleks inisiasi eukariotik mengenali tudung 7-metilguanosin di ujung 5′ mRNA. Protein pengikat tudung (CBP) dan beberapa IF lainnya membantu pergerakan ribosom ke tudung 5′. Setelah berada di tudung, kompleks inisiasi menelusuri mRNA dari arah 5′ ke 3′, mencari kodon awal AUG. Banyak mRNA eukariotik diterjemahkan dari AUG pertama, tetapi ini tidak selalu terjadi. Menurut aturan Kozak , nukleotida di sekitar AUG menunjukkan apakah itu kodon awal yang benar. Aturan Kozak menyatakan bahwa sekuens konsensus berikut harus muncul di sekitar AUG gen vertebrata: 5′-gccRccAUGG-3′. Huruf R (untuk purin) menunjukkan situs yang dapat berupa A atau G, tetapi tidak dapat berupa C atau U. Pada dasarnya, semakin dekat urutan tersebut dengan konsensus ini, semakin tinggi efisiensi translasi.
Setelah kodon AUG yang sesuai teridentifikasi, protein lain dan CBP akan berpisah, dan subunit 60S akan berikatan dengan kompleks Met-tRNAi, mRNA, dan subunit 40S. Langkah ini menyelesaikan inisiasi translasi pada eukariota.
Translasi, Pemanjangan, dan Terminasi
Pada prokariota dan eukariota, dasar-dasar elongasi adalah sama, jadi kita akan meninjau elongasi dari perspektif E. coli . Subunit ribosom 50S E. coli terdiri dari tiga kompartemen: situs A (aminoasil) mengikat tRNA aminoasil bermuatan yang masuk. Situs P (peptidil) mengikat tRNA bermuatan yang membawa asam amino yang telah membentuk ikatan peptida dengan rantai polipeptida yang sedang tumbuh tetapi belum terlepas dari tRNA yang sesuai. Situs E (keluar) melepaskan tRNA yang terlepas sehingga dapat diisi ulang dengan asam amino bebas. Ada satu pengecualian untuk jalur perakitan tRNA ini: pada E. coli , mampu memasuki situs P secara langsung tanpa terlebih dahulu memasuki situs A. Demikian pula, Met-tRNAi eukariotik, dengan bantuan protein lain dari kompleks inisiasi, mengikat langsung ke situs P (Gambar 1). Dalam kedua kasus, ini menciptakan kompleks inisiasi dengan situs A bebas yang siap menerima tRNA yang sesuai dengan kodon pertama setelah AUG
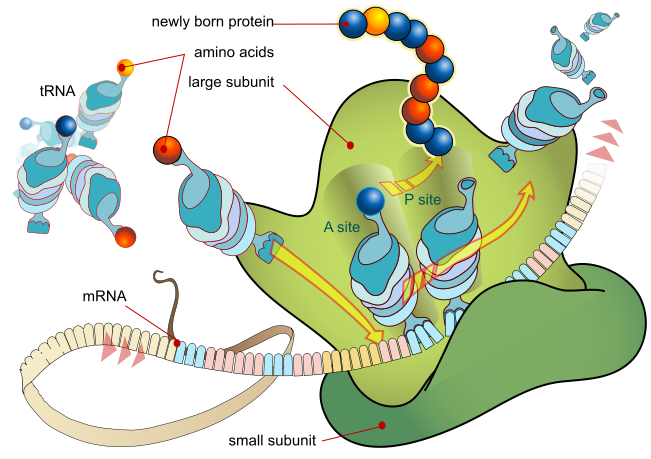
Gambar 1. Translasi mRNA ribosom
Selama elongasi translasi, templat mRNA memberikan spesifisitas. Saat ribosom bergerak di sepanjang mRNA, setiap kodon mRNA berada pada posisi yang tepat, dan pengikatan spesifik dengan antikodon tRNA bermuatan yang sesuai dipastikan terjadi. Jika mRNA tidak ada dalam kompleks elongasi, ribosom akan mengikat tRNA secara nonspesifik.
Elongasi berlangsung dengan tRNA bermuatan memasuki situs A dan kemudian bergeser ke situs P diikuti oleh situs E dengan setiap “langkah” kodon tunggal ribosom. Langkah-langkah ribosom diinduksi oleh perubahan konformasi yang memajukan ribosom sebanyak tiga basa ke arah 3′. Energi untuk setiap langkah ribosom disumbangkan oleh faktor elongasi yang menghidrolisis GTP. Ikatan peptida terbentuk antara gugus amino dari asam amino yang terikat pada tRNA situs A dan gugus karboksil dari asam amino yang terikat pada tRNA situs P. Pembentukan setiap ikatan peptida dikatalisis oleh peptidyl transferase , enzim berbasis RNA yang terintegrasi ke dalam subunit ribosom 50S. Energi untuk setiap pembentukan ikatan peptida berasal dari hidrolisis GTP, yang dikatalisis oleh faktor elongasi terpisah. Asam amino yang terikat pada tRNA situs P juga terhubung ke rantai polipeptida yang sedang tumbuh. Saat ribosom melangkah melintasi mRNA, tRNA di situs P sebelumnya memasuki situs E, terlepas dari asam amino, dan dikeluarkan (Gambar 2). Hebatnya, aparatus translasi E. coli hanya membutuhkan 0,05 detik untuk menambahkan setiap asam amino, yang berarti bahwa protein 200 asam amino dapat diterjemahkan hanya dalam 10 detik.
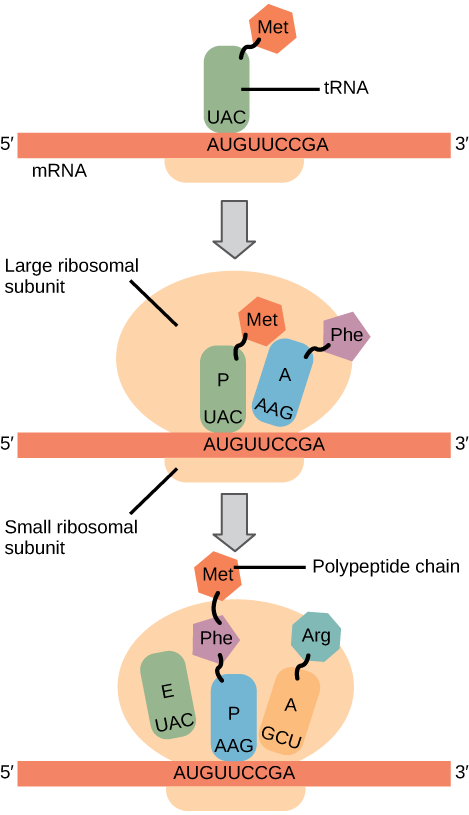
Gambar 2. Translasi dimulai ketika antikodon tRNA inisiator mengenali kodon pada mRNA. Subunit ribosom besar bergabung dengan subunit kecil, dan tRNA kedua direkrut. Saat mRNA bergerak relatif terhadap ribosom, rantai polipeptida terbentuk. Masuknya faktor pelepasan ke situs A mengakhiri translasi dan komponen-komponen tersebut berpisah.
Penghentian translasi terjadi ketika kodon nonsens (UAA, UAG, atau UGA) ditemukan. Setelah sejajar dengan situs A, kodon nonsens ini dikenali oleh faktor pelepasan pada prokariota dan eukariota yang menginstruksikan peptidyl transferase untuk menambahkan molekul air ke ujung karboksil asam amino situs P. Reaksi ini memaksa asam amino situs P untuk terlepas dari tRNA-nya, dan protein yang baru dibuat dilepaskan. Subunit ribosom kecil dan besar berpisah dari mRNA dan satu sama lain; mereka direkrut hampir segera ke dalam kompleks inisiasi translasi lainnya. Setelah banyak ribosom menyelesaikan translasi, mRNA didegradasi sehingga nukleotida dapat digunakan kembali dalam reaksi transkripsi lain.
Pemrosesan tRNA dan rRNA
tRNA dan rRNA adalah molekul struktural yang berperan dalam sintesis protein; namun, RNA ini sendiri tidak diterjemahkan. Pra-rRNA ditranskripsikan, diproses, dan dirakit menjadi ribosom di nukleolus. Pra-tRNA ditranskripsikan dan diproses di nukleus dan kemudian dilepaskan ke sitoplasma di mana mereka dihubungkan dengan asam amino bebas untuk sintesis protein.
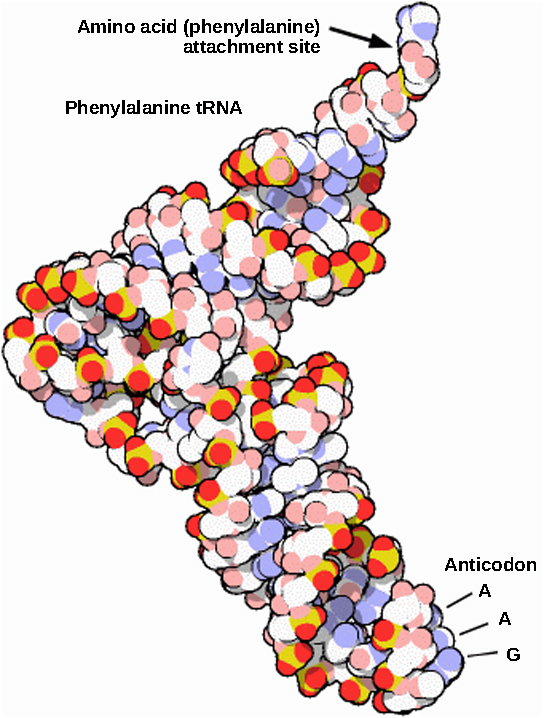
Gambar 3. Ini adalah model pengisian ruang dari molekul tRNA yang menambahkan asam amino fenilalanin ke rantai polipeptida yang sedang tumbuh. Antikodon AAG mengikat Kodon UUC pada mRNA. Asam amino fenilalanin terikat pada ujung tRNA lainnya.
Sebagian besar tRNA dan rRNA pada eukariota dan prokariota pertama kali ditranskripsikan sebagai molekul prekursor panjang yang mencakup beberapa rRNA atau tRNA. Enzim kemudian membelah prekursor menjadi subunit yang sesuai dengan setiap RNA struktural. Beberapa basa pre-rRNA dimetilasi; yaitu, gugus –CH3
( gugus fungsional metil) ditambahkan untuk stabilitas. Molekul pre-tRNA juga mengalami metilasi. Seperti halnya pre-mRNA, eksisi subunit terjadi pada pre-RNA eukariotik yang ditakdirkan untuk menjadi tRNA atau rRNA.
rRNA matang membentuk sekitar 50 persen dari setiap ribosom. Beberapa molekul RNA ribosom bersifat struktural murni, sedangkan yang lain memiliki aktivitas katalitik atau pengikatan. tRNA matang mengambil struktur tiga dimensi melalui ikatan hidrogen intramolekuler untuk memposisikan situs pengikatan asam amino di satu ujung dan antikodon di ujung lainnya (Gambar 3).
Antikodon adalah urutan tiga nukleotida dalam tRNA yang berinteraksi dengan kodon mRNA melalui pasangan basa komplementer.
Ribosom
Sintesis protein mengonsumsi lebih banyak energi sel daripada proses metabolisme lainnya. Sebagai imbalannya, protein menyumbang massa lebih besar daripada komponen organisme hidup lainnya (kecuali air), dan protein menjalankan hampir semua fungsi sel. Proses translasi, atau sintesis protein, melibatkan penguraian pesan mRNA menjadi produk polipeptida. Asam amino dirangkai secara kovalen oleh ikatan peptida yang saling terkait dengan panjang mulai dari sekitar 50 residu asam amino hingga lebih dari 1.000. Setiap asam amino memiliki gugus amino (NH₂ ) dan gugus karboksil (COOH). Polipeptida terbentuk ketika gugus amino dari satu asam amino membentuk ikatan amida (yaitu, peptida) dengan gugus karboksil dari asam amino lainnya (Gambar 4). Reaksi ini dikatalisis oleh ribosom dan menghasilkan satu molekul air.
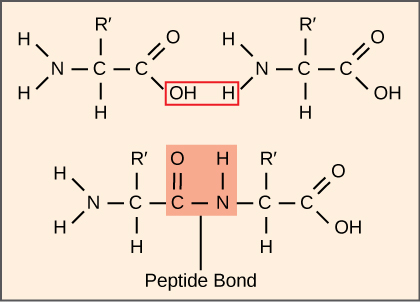
Gambar 4. Ikatan peptida menghubungkan ujung karboksil dari satu asam amino dengan ujung amino dari asam amino lainnya, mengeluarkan satu molekul air. Untuk penyederhanaan pada gambar ini, hanya gugus fungsional yang terlibat dalam ikatan peptida yang ditampilkan. Penamaan R dan R′ mengacu pada bagian struktur asam amino lainnya
Mesin Sintesis Protein
Selain templat mRNA, banyak molekul dan makromolekul berkontribusi pada proses translasi. Komposisi setiap komponen dapat bervariasi antar spesies; misalnya, ribosom dapat terdiri dari jumlah rRNA dan polipeptida yang berbeda tergantung pada organisme. Namun, struktur dan fungsi umum mesin sintesis protein dapat dibandingkan dari bakteri hingga sel manusia. Translasi membutuhkan masukan berupa templat mRNA, ribosom, tRNA, dan berbagai faktor enzimatik.
Ribosom
Bahkan sebelum mRNA diterjemahkan, sel harus menginvestasikan energi untuk membangun setiap ribosomnya. Pada E. coli , terdapat antara 10.000 dan 70.000 ribosom di setiap sel pada waktu tertentu. Ribosom adalah makromolekul kompleks yang terdiri dari rRNA struktural dan katalitik, serta banyak polipeptida yang berbeda. Pada eukariota, nukleolus sepenuhnya terspesialisasi untuk sintesis dan perakitan rRNA
Ribosom terdapat di sitoplasma pada prokariota dan di sitoplasma serta retikulum endoplasma kasar pada eukariota. Mitokondria dan kloroplas juga memiliki ribosomnya sendiri di matriks dan stroma, yang lebih mirip dengan ribosom prokariotik (dan memiliki sensitivitas obat yang serupa) daripada ribosom yang berada tepat di luar membran luarnya di sitoplasma. Ribosom berdisosiasi menjadi subunit besar dan kecil ketika tidak mensintesis protein dan bergabung kembali selama inisiasi translasi. Pada E. coli , subunit kecil digambarkan sebagai 30S, dan subunit besar adalah 50S, dengan total 70S (ingat bahwa satuan Svedberg tidak bersifat aditif). Ribosom mamalia memiliki subunit kecil 40S dan subunit besar 60S, dengan total 80S. Subunit kecil bertanggung jawab untuk mengikat templat mRNA, sedangkan subunit besar secara berurutan mengikat tRNA. Setiap molekul mRNA diterjemahkan secara simultan oleh banyak ribosom, semuanya mensintesis protein dalam arah yang sama: membaca mRNA dari 5′ ke 3′ dan mensintesis polipeptida dari ujung N ke ujung C. Struktur mRNA/poli-ribosom yang lengkap disebut polisom .
tRNA
tRNA adalah molekul RNA struktural yang ditranskripsikan dari gen oleh RNA polimerase III. Tergantung pada spesiesnya, terdapat 40 hingga 60 jenis tRNA di sitoplasma. Berfungsi sebagai adaptor, tRNA spesifik mengikat sekuens pada templat mRNA dan menambahkan asam amino yang sesuai ke rantai polipeptida. Oleh karena itu, tRNA adalah molekul yang sebenarnya “menerjemahkan” bahasa RNA ke dalam bahasa protein
Dari 64 kemungkinan kodon mRNA—atau kombinasi triplet A, U, G, dan C—tiga di antaranya menentukan penghentian sintesis protein dan 61 menentukan penambahan asam amino ke rantai polipeptida. Dari 61 kodon tersebut, satu kodon (AUG) juga mengkodekan inisiasi translasi. Setiap antikodon tRNA dapat berpasangan dengan salah satu kodon mRNA dan menambahkan asam amino atau menghentikan translasi, sesuai dengan kode genetik. Misalnya, jika sekuens CUA muncul pada templat mRNA dalam kerangka baca yang tepat, ia akan mengikat tRNA yang mengekspresikan sekuens komplementer, GAU, yang akan dihubungkan dengan asam amino leusin.
Sebagai molekul adaptor translasi, sungguh mengejutkan bahwa tRNA dapat memuat begitu banyak spesifisitas dalam paket yang begitu kecil. Perhatikan bahwa tRNA perlu berinteraksi dengan tiga faktor:
- Mereka harus dikenali oleh aminoasil sintetase yang tepat.
- Mereka harus dikenali oleh ribosom.
- Mereka harus berikatan dengan sekuens yang tepat dalam mRNA.
Sintetase Aminoasil tRNA
Proses sintesis pra-tRNA oleh RNA polimerase III hanya menciptakan bagian RNA dari molekul adaptor. Asam amino yang sesuai harus ditambahkan kemudian, setelah tRNA diproses dan diekspor ke sitoplasma. Melalui proses “pengisian” tRNA, setiap molekul tRNA dihubungkan dengan asam amino yang tepat oleh sekelompok enzim yang disebut sintetase aminoasil tRNA. Setidaknya satu jenis sintetase aminoasil tRNA ada untuk masing-masing dari 20 asam amino; jumlah pasti sintetase aminoasil tRNA bervariasi menurut spesies. Enzim-enzim ini pertama-tama mengikat dan menghidrolisis ATP untuk mengkatalisis ikatan berenergi tinggi antara asam amino dan adenosin monofosfat (AMP); molekul pirofosfat dikeluarkan dalam reaksi ini. Asam amino yang diaktifkan kemudian ditransfer ke tRNA, dan AMP dilepaskan
Kodon
Mengingat perbedaan jumlah “huruf” dalam “alfabet” mRNA dan protein, para ilmuwan berteori bahwa kombinasi nukleotida sesuai dengan asam amino tunggal. Pasangan nukleotida tidak akan cukup untuk menentukan setiap asam amino karena hanya ada 16 kemungkinan kombinasi dua nukleotida (42). Sebaliknya, ada 64 kemungkinan triplet nukleotida (43), yang jauh lebih banyak daripada jumlah asam amino. Para ilmuwan berteori bahwa asam amino dikodekan oleh triplet nukleotida dan bahwa kode genetik bersifat degeneratif . Dengan kata lain, asam amino tertentu dapat dikodekan oleh lebih dari satu triplet nukleotida. Hal ini kemudian dikonfirmasi secara eksperimental; Francis Crick dan Sydney Brenner menggunakan mutagen kimia proflavin untuk memasukkan satu, dua, atau tiga nukleotida ke dalam gen virus. Ketika satu atau dua nukleotida dimasukkan, sintesis protein sepenuhnya terhenti. Ketika tiga nukleotida dimasukkan, protein disintesis dan berfungsi. Hal ini menunjukkan bahwa tiga nukleotida menentukan setiap asam amino. Tiga nukleotida ini disebut kodon . Penyisipan satu atau dua nukleotida sepenuhnya mengubah kerangka baca triplet, sehingga mengubah pesan untuk setiap asam amino berikutnya (Gambar 5). Meskipun penyisipan tiga nukleotida menyebabkan asam amino tambahan dimasukkan selama translasi, integritas bagian protein lainnya tetap terjaga.
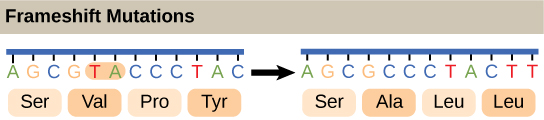
Gambar 5. Penghapusan dua nukleotida menggeser kerangka baca mRNA dan mengubah seluruh pesan protein, menciptakan protein yang tidak berfungsi atau menghentikan sintesis protein sama sekali.
Para ilmuwan dengan susah payah memecahkan kode genetik dengan menerjemahkan mRNA sintetis secara in vitro dan mengurutkan protein yang dihasilkan (Gambar 6).
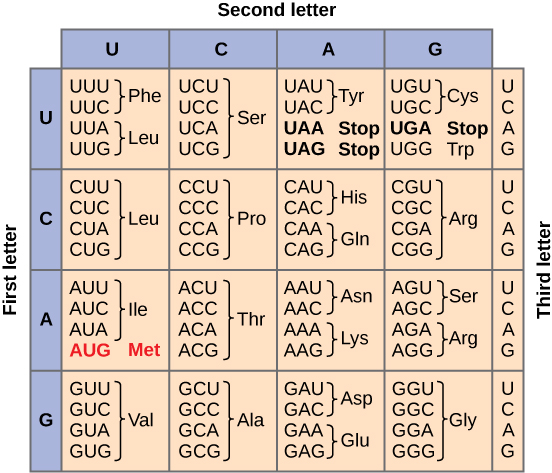
Gambar 6. Gambar ini menunjukkan kode genetik untuk menerjemahkan setiap triplet nukleotida dalam mRNA menjadi asam amino atau sinyal terminasi dalam protein yang baru terbentuk. (kredit: modifikasi karya NIH)
Selain menginstruksikan penambahan asam amino spesifik ke rantai polipeptida, tiga dari 64 kodon mengakhiri sintesis protein dan melepaskan polipeptida dari mesin translasi. Triplet ini disebut kodon nonsens, atau kodon berhenti. Kodon lain, AUG, juga memiliki fungsi khusus. Selain menentukan asam amino metionin, ia juga berfungsi sebagai kodon awal untuk memulai translasi. Kerangka baca untuk translasi ditentukan oleh kodon awal AUG di dekat ujung 5′ mRNA.
Kode genetik bersifat universal. Dengan beberapa pengecualian, hampir semua spesies menggunakan kode genetik yang sama untuk sintesis protein. Konservasi kodon berarti bahwa mRNA murni yang mengkode protein globin pada kuda dapat ditransfer ke sel tulip, dan tulip akan mensintesis globin kuda. Fakta bahwa hanya ada satu kode genetik merupakan bukti kuat bahwa semua kehidupan di Bumi memiliki asal usul yang sama, terutama mengingat ada sekitar 1084 kemungkinan kombinasi dari 20 asam amino dan 64 kodon triplet.
Degenerasi diyakini sebagai mekanisme seluler untuk mengurangi dampak negatif mutasi acak. Kodon yang menentukan asam amino yang sama biasanya hanya berbeda satu nukleotida. Selain itu, asam amino dengan rantai samping yang secara kimiawi serupa dikodekan oleh kodon yang serupa. Nuansa kode genetik ini memastikan bahwa mutasi substitusi satu nukleotida dapat menentukan asam amino yang sama tetapi tidak berpengaruh, atau menentukan asam amino yang serupa, mencegah protein menjadi sepenuhnya tidak berfungsi.
Modifikasi Protein
Selama dan setelah translasi, asam amino individual dapat dimodifikasi secara kimiawi, sekuens sinyal dapat ditambahkan, dan protein baru “melipat” menjadi struktur tiga dimensi yang berbeda sebagai hasil dari interaksi intramolekuler. Sekuens sinyal adalah ekor pendek asam amino yang mengarahkan protein ke kompartemen seluler tertentu. Sekuens di ujung amino atau ujung karboksil protein ini dapat dianggap sebagai “tiket kereta” protein ke tujuan akhirnya. Faktor seluler lainnya mengenali setiap sekuens sinyal dan membantu mengangkut protein dari sitoplasma ke kompartemen yang tepat. Misalnya, sekuens spesifik di ujung amino akan mengarahkan protein ke mitokondria atau kloroplas (pada tumbuhan). Setelah protein mencapai tujuan selulernya, sekuens sinyal biasanya dipotong
Banyak protein melipat secara spontan, tetapi beberapa protein membutuhkan molekul pembantu, yang disebut chaperone, untuk mencegahnya menggumpal selama proses pelipatan yang rumit. Bahkan jika suatu protein ditentukan dengan benar oleh mRNA yang sesuai, protein tersebut dapat mengambil bentuk yang sama sekali tidak berfungsi jika kondisi suhu atau pH yang tidak normal mencegahnya melipat dengan benar.
Modifikasi Kimia, Aktivitas Protein, dan Umur Panjang
Protein dapat dimodifikasi secara kimiawi dengan penambahan gugus-gugus termasuk gugus metil, fosfat, asetil, dan ubiquitin. Penambahan atau penghapusan gugus-gugus ini dari protein mengatur aktivitasnya atau lamanya protein tersebut berada di dalam sel. Terkadang modifikasi ini dapat mengatur di mana protein tersebut ditemukan di dalam sel—misalnya, di nukleus, sitoplasma, atau menempel pada membran plasma.
Modifikasi kimia terjadi sebagai respons terhadap rangsangan eksternal seperti stres, kekurangan nutrisi, panas, atau paparan sinar ultraviolet. Perubahan ini dapat mengubah aksesibilitas epigenetik, transkripsi, stabilitas mRNA, atau translasi—semuanya mengakibatkan perubahan ekspresi berbagai gen. Ini adalah cara yang efisien bagi sel untuk dengan cepat mengubah kadar protein spesifik sebagai respons terhadap lingkungan. Karena protein terlibat dalam setiap tahap regulasi gen, fosforilasi protein (tergantung pada protein yang dimodifikasi) dapat mengubah aksesibilitas ke kromosom, dapat mengubah translasi (dengan mengubah pengikatan atau fungsi faktor transkripsi), dapat mengubah perpindahan nuklir (dengan memengaruhi modifikasi pada kompleks pori nuklir), dapat mengubah stabilitas RNA (dengan mengikat atau tidak mengikat RNA untuk mengatur stabilitasnya), dapat memodifikasi translasi (meningkatkan atau menurunkan), atau dapat mengubah modifikasi pasca-translasi (menambah atau menghilangkan fosfat atau modifikasi kimia lainnya).
Penambahan gugus ubiquitin pada protein menandai protein tersebut untuk degradasi. Ubiquitin bertindak seperti bendera yang menunjukkan bahwa masa hidup protein telah selesai. Protein-protein ini dipindahkan ke proteasom , organel yang berfungsi untuk membuang protein, untuk didegradasi (Gambar 7). Oleh karena itu, salah satu cara untuk mengontrol ekspresi gen adalah dengan mengubah umur protein.
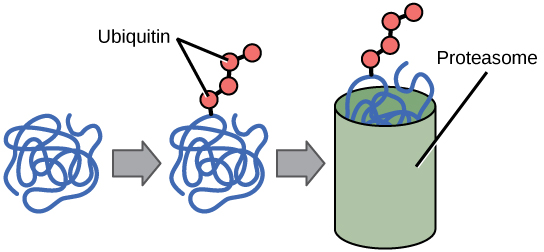
Gambar 7. Protein dengan penanda ubiquitin ditandai untuk degradasi di dalam proteasom.
Sumber :
- Biology. Provided by: OpenStax CNX. Located at: http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8. License: CC BY: Attribution. License Terms: Download for free at http://cnx.org/contents/185cbf87-c72e-48f5-b51e-f14f21b5eabd@10.8
- Introduction to Translation. Authored by: Shelli Carter and Lumen Learning. Provided by: Lumen Learning. License: CC BY: Attribution
- https://courses.lumenlearning.com/suny-wmopen-biology1/chapter/translation/
Ringkasan Materi:
- DNA adalah materi genetik berbentuk heliks ganda yang menyimpan informasi biologis melalui urutan basa A, T, C, dan G.
- Genetik adalah proses pewarisan informasi DNA dari orang tua kepada keturunannya.
- Genom adalah keseluruhan DNA dalam suatu organisme yang menyimpan seluruh instruksi kehidupan.
- Kromosom adalah struktur tempat DNA dikemas yang berjumlah 46 pada manusia dan diwariskan dari kedua orang tua.
- Gen adalah segmen DNA yang mengandung instruksi untuk membentuk protein.
- Protein adalah molekul hasil ekspresi gen yang berperan dalam struktur dan fungsi tubuh.
- Alel adalah variasi bentuk suatu gen yang menyebabkan perbedaan sifat antar individu.
- Sel adalah unit dasar kehidupan yang mengandung DNA di dalam nukleusnya.
- Ekspresi gen adalah proses penggunaan informasi gen untuk menghasilkan protein.
- Metilasi DNA adalah mekanisme epigenetik yang mengatur aktivitas gen tanpa mengubah urutan DNA.
- Perubahan genetik (mutasi/varian) adalah perubahan urutan basa DNA yang dapat memengaruhi fungsi protein.
- Copy Number Variation (CNV) adalah variasi jumlah salinan segmen DNA yang dapat memengaruhi keragaman genetik.
- Faktor lingkungan adalah paparan eksternal yang dapat memengaruhi cara kerja gen.
- Replikasi DNA adalah proses penggandaan DNA sebelum pembelahan sel agar informasi genetik dapat diwariskan secara akurat.
- Transkripsi adalah proses penyalinan informasi dari DNA menjadi RNA sebagai langkah awal sintesis protein.
- Translasi adalah proses penerjemahan kode RNA menjadi rantai asam amino yang membentuk protein.
- Kode genetik adalah sistem aturan yang menentukan bagaimana urutan basa pada RNA diterjemahkan menjadi asam amino.
Simak Video Pembelajaran dibawah ini !