Dalam organisme multiseluler, pembelahan sel bukan sekadar proses reproduksi sel, melainkan mekanisme yang diatur secara ketat oleh sinyal genetik.
Pada Sel Imun: Pembelahan bersifat episodik. Sel hanya membelah secara masif saat ada aktivasi antigen (misal: infeksi), lalu berhenti atau mati setelah ancaman hilang.
Pada Sel Kanker: Pembelahan bersifat otonom. Terjadi kegagalan pada “sakelar” genetik yang seharusnya menghentikan siklus sel.
1. Dasar Genetik Kontrol Siklus Sel
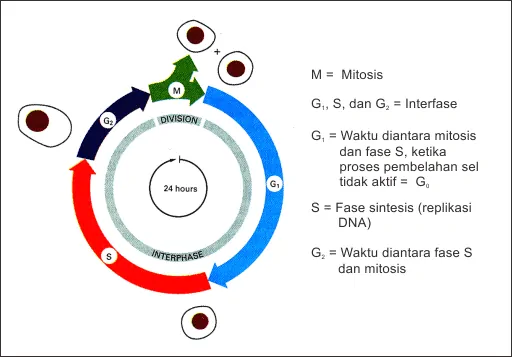
Gambar 1 : Siklus Sel
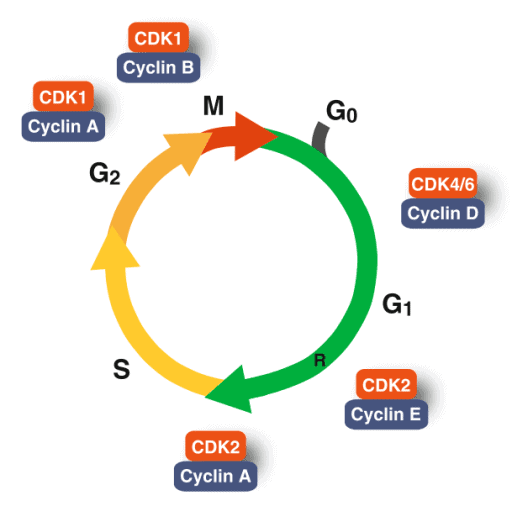
Dasar genetik kontrol siklus sel melibatkan interaksi kompleks antara protein regulator, terutama Siklin (cyclin) dan Cyclin-Dependent Kinases (CDK), yang mengatur kemajuan sel melalui fase G1, S, G2, dan M. Sistem ini memastikan replikasi DNA yang akurat melalui checkpoint (titik pemeriksaan) genetik, yang dipicu oleh sinyal internal/eksternal untuk mencegah kanker atau kerusakan DNA.
Siklus sel dikendalikan oleh interaksi antara protein yang diproduksi oleh dua kelompok gen utama:
Kelompok Gen
Fungsi Normal
Perubahan pada Kanker
Proto-onkogen
Mendorong pembelahan sel yang sehat (seperti pedal gas).
Bermutasi menjadi Onkogen (gas terkunci/aktif terus menerus).
Tumor Suppressor Genes (TSG)
Menghambat pembelahan dan memperbaiki DNA (seperti rem).
Mengalami Inaktivasi (rem blong).
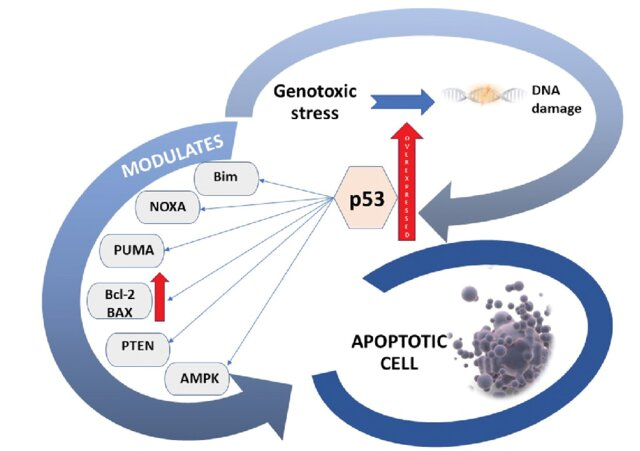
Contoh Penting: Gen p53 (The Guardian of the Genome). Jika DNA rusak, p53 akan menghentikan siklus sel di fase G1 untuk perbaikan atau memicu apoptosis jika kerusakan terlalu parah.
2. Kontrol Genetik pada Sel Imun
Sel imun (khususnya Limfosit T dan B) memiliki keunikan genetik yang tidak dimiliki sel lain:
Rekombinasi V(D)J: Proses pemotongan dan penyambungan gen secara sengaja untuk menciptakan keragaman antibodi/reseptor.
Proliferasi Klonal: Ketika antigen terdeteksi, gen pengatur siklus sel memicu pembelahan yang sangat cepat (ekspansi).
Checkpoints Imun: Adanya gen pengontrol seperti CTLA-4 dan PD-1 yang berfungsi sebagai “rem molekuler” untuk mencegah limfosit membelah tanpa kendali yang bisa menyebabkan autoimun.
3. Molekul Pengatur: Cyclin dan CDK
Secara molekuler, pembelahan sel dikendalikan oleh pasangan protein:
Cyclin: Protein yang konsentrasinya naik-turun sesuai fase siklus sel.
CDK (Cyclin-Dependent Kinases): Enzim yang aktif hanya saat berikatan dengan Cyclin.
Pada sel kanker, sering kali terjadi overekspresi gen Cyclin atau mutasi pada inhibitor CDK, sehingga sel terus melewati fase pemeriksaan (checkpoints) tanpa verifikasi keamanan DNA
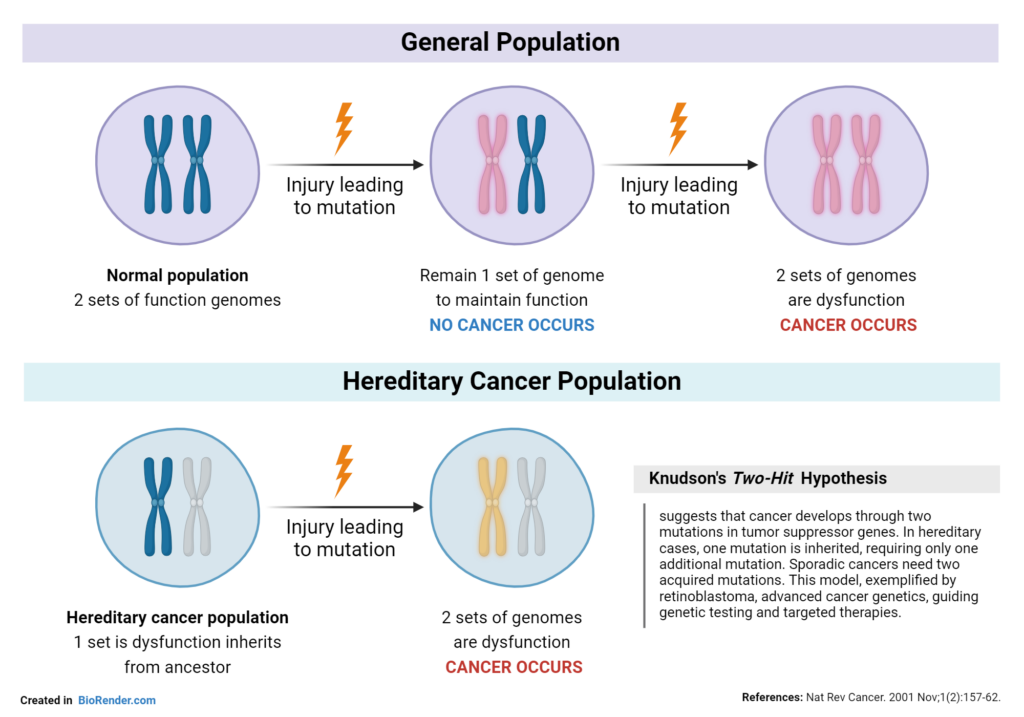
4. Teori “Two-Hit Hypothesis” (Knudson)
Mahasiswa perlu memahami mengapa kanker seringkali memerlukan waktu lama untuk berkembang:
Hit 1: Mutasi pertama pada satu alel gen penekan tumor (bisa keturunan atau sporadis).
Hit 2: Mutasi pada alel kedua. Setelah kedua “rem” ini rusak, kontrol pembelahan hilang sepenuhnya, memicu transformasi sel normal menjadi sel ganas.
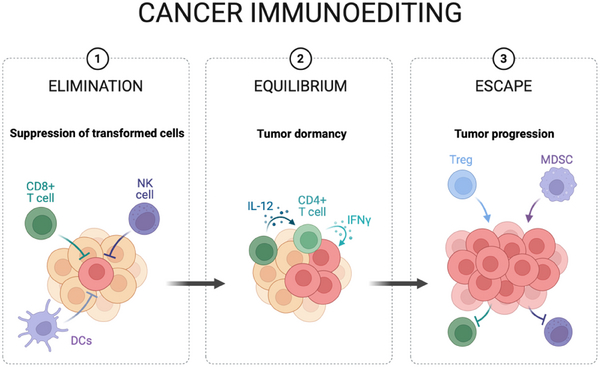
5. Hubungan Imun dan Kanker: Immunoediting
Sistem imun sebenarnya adalah “polisi genetik” yang memantau sel-sel yang pembelahannya mulai tidak normal. Kanker terjadi jika:
Escape: Sel kanker bermutasi sehingga tidak lagi mengekspresikan antigen yang bisa dikenali sel imun.
Exhaustion: Sel imun menjadi “lelah” karena sinyal pembelahan kanker yang terlalu persisten
Sel Imun dan Kanker
Pengendalian pembelahan sel imun sangat mendasar bagi kanker, bertindak sebagai pengatur penting pertumbuhan tumor.Sistem kekebalan tubuh, khususnya sel Natural Killer (NK) dan limfosit T sitotoksik, memantau dan menghancurkan sel-sel yang menunjukkan pembelahan yang tidak terkontrol. Namun, sel kanker sering kali lolos dari deteksi ini dengan membajak proses pengaturan normal, seperti pos pemeriksaan kekebalan dan kontrol siklus sel, yang menyebabkan penghindaran kekebalan.
Mekanisme Pengendalian Kekebalan Tubuh
Pengawasan imun: Tubuh terus-menerus memantau sel-sel abnormal dengan DNA yang rusak, menggunakan sel darah putih untuk menghancurkannya.
Pos Pemeriksaan Imun: Reseptor seperti PD-L1 dan CTLA-4 digunakan oleh sel imun untuk menjaga homeostasis dan mencegah autoimunitas; namun, sel kanker mengekspresikan reseptor ini secara berlebihan untuk menginduksi “kelelahan” sel T dan menghindari penghancuran.
Regulasi Sitokin: Sel imun inflamasi (misalnya, sel T) dapat menghambat pertumbuhan tumor, tetapi makrofag yang terkait dengan tumor terkadang dapat memicu pertumbuhan tumor.
Bagaimana Sel Kanker Menghindari Kontrol Kekebalan Tubuh
Penekanan Sistem Kekebalan Tubuh: Tumor menciptakan lingkungan imunosupresif dengan mensekresikan zat-zat seperti TGF-β dan merekrut sel T regulator.
Resistensi Mitosis: Sel kanker dalam fase mitosis (pembelahan aktif) mengekspresikan protein penghambat kekebalan tubuh seperti HLA-G dan PD-L1 dalam kadar yang lebih tinggi, sehingga membuatnya lebih resisten terhadap serangan kekebalan dibandingkan dengan sel dalam fase istirahat.
Mutasi Genetik: Sel kanker memutasi gen-gen kunci, seperti gen-gen dalam jalur presentasi antigen MHC kelas I, sehingga menjadi tidak terlihat oleh sel T CD8+.
Strategi Terapi
Imunoterapi: Teknik-teknik seperti terapi sel T CAR, antibodi monoklonal, dan penghambat titik kontrol imun (misalnya, anti-PD-L1) dirancang untuk “mengungkap” sel kanker dan meningkatkan sistem kekebalan tubuh.
Inhibitor Siklus Sel: Obat-obatan yang menghambat siklus sel, khususnya inhibitor CDK4/6, dapat menurunkan ekspresi PD-L1 dengan menjaga sel tetap berada dalam fase G1, sehingga meningkatkan infiltrasi sel imun dan meningkatkan efektivitas pengobatan lainnya.
Peringatan tentang Penghambatan Mitosis: Meskipun penghambat mitosis (misalnya, paclitaxel) umum digunakan, beberapa penelitian menunjukkan bahwa menginduksi penghambatan mitosis total justru dapat meningkatkan resistensi terhadap serangan imun.
Tabel Ringkasan Interaksi Sel Imun-Kanker
Komponen
Fungsi dalam Pengendalian/Perkembangan Kanker
Sel T Sitotoksik
Membunuh sel tumor melalui pengenalan antigen asing
Sel Pembunuh Alami (NK)
Pertahanan lini pertama, penghancuran sel abnormal secara bawaan.
PD-L1/HLA-G
Titik-titik pemeriksaan yang dieksploitasi oleh kanker untuk menghindari serangan sel T.
Inhibitor CDK4/6
Meningkatkan infiltrasi imun dengan menghentikan siklus sel di fase G1.
Lingkungan Mikro Tumor
Dapat dimanipulasi oleh kanker untuk bersifat imunosupresif
Keseimbangan antara laju pembelahan sel dan pengawasan imun menentukan apakah tumor diberantas atau berkembang.
A. Sel Imun
Sel Imun adalah objek studi yang luar biasa karena mereka adalah satu-satunya sel somatik dalam tubuh manusia yang secara rutin melakukan pengaturan ulang genom (DNA rearrangement) sebagai bagian dari fungsi normalnya.
1.Keunikan Genetik: Rekombinasi Somatik
Berbeda dengan sel kulit atau sel otot yang menjaga urutan DNA-nya tetap statis, sel imun (limfosit B dan T) melakukan modifikasi DNA yang dramatis:
Implikasi Kontrol: Proses pemotongan DNA ini sangat berisiko. Jika terjadi kesalahan penyambungan (translokasi), gen yang seharusnya mengontrol pembelahan (seperti MYC) bisa berpindah ke dekat promotor gen imun yang sangat aktif, memicu pembelahan tak terkendali (Limfoma).
V(D)J Recombination: Sel imun memotong dan menyambung kembali gen mereka sendiri untuk menghasilkan jutaan variasi reseptor antigen.
2. Ekspansi Klonal (Pembelahan Terprogram) Sel imun memiliki kontrol siklus sel yang sangat responsif:
Terminasi: Setelah infeksi mereda, gen pro-apoptosis (seperti Bax atau Bak) diaktifkan untuk memangkas jumlah sel imun yang berlebih, menyisakan hanya sel memori.
Fase Quiescent (G0): Sebagian besar sel imun berada dalam fase istirahat.
Aktivasi: Ketika reseptor mengenali antigen, sel menerima sinyal genetik untuk masuk ke siklus sel secara masif. Ini adalah contoh pembelahan sel yang paling cepat pada mamalia dewasa.
3. Checkpoints: Penjaga Keamanan
Sistem imun memiliki “checkpoints” genetik untuk memastikan mereka tidak menyerang tubuh sendiri atau membelah tanpa henti:
CTLA-4 & PD-1: Ini adalah protein permukaan yang berfungsi sebagai “rem”. Secara genetik, ekspresi protein ini meningkat setelah sel imun aktif beberapa lama untuk mencegah kerusakan jaringan.
Koneksi Kanker: Sel kanker sering kali “mencuri” mekanisme ini dengan mengekspresikan ligan (PD-L1) yang menekan sel imun, membuat sel imun “tertidur” dan gagal mengontrol tumor.
B. Sel Kanker
Dalam studi genetika, sel kanker didefinisikan sebagai sel yang mengalami akumulasi mutasi sehingga kehilangan homeostasis pembelahan. Jika sel imun adalah “tentara yang disiplin”, maka sel kanker adalah “pemberontak” yang mengabaikan semua regulasi molekuler.
Berikut adalah pilar utama materi mengenai genetika sel kanker:
1. Hallmarks of Cancer (Sifat Dasar Genetik)
Menurut konsep Hanahan dan Weinberg, sel kanker memiliki kemampuan genetik untuk:
Self-sufficiency: Memproduksi sinyal pertumbuhan sendiri tanpa perintah tubuh.
Insensitivity: Mengabaikan sinyal anti-proliferasi (rem biologis).
Evading Apoptosis: Mematikan jalur bunuh diri sel meski DNA rusak parah.
Limitless Replication: Mengekspresikan enzim Telomerase agar kromosom tidak memendek, sehingga sel bisa membelah selamanya (immortal).
2. Akumulasi Mutasi: “Driver” vs “Passenger”
Tidak semua mutasi menyebabkan kanker. Mahasiswa harus membedakan:
Driver Mutations: Mutasi pada gen kunci (seperti TP53, RAS, APC) yang langsung mendorong transformasi keganasan.
Passenger Mutations: Mutasi “penumpang” yang terjadi akibat instabilitas genom tetapi tidak berkontribusi langsung pada progresivitas kanker.
3. Instabilitas Genomik
Sel kanker sering kehilangan kemampuan DNA Repair. Hal ini menyebabkan:
Aneuploidi: Jumlah kromosom yang tidak normal (misal: 47 atau 92 kromosom).
Translokasi: Potongan kromosom berpindah ke tempat yang salah, sering kali menggabungkan dua gen menjadi satu protein baru yang berbahaya (contoh: Kromosom Philadelphia pada Leukemia).
4. Epigenetik Kanker
Kontrol pembelahan tidak hanya rusak karena perubahan urutan DNA, tetapi juga perubahan ekspresi gen:
Hipermetilasi Promotor: Menempelnya gugus metil pada promotor gen penekan tumor (Tumor Suppressor Gene), sehingga gen tersebut “bungkam” meskipun urutan DNA-nya normal.
5. Interaksi dengan Sel Imun (Immune Evasion)
Sel kanker melakukan “penyamaran genetik” agar tidak dihancurkan oleh sel imun melalui:
Downregulation MHC: Menghilangkan “papan pengenal” sel sehingga sel T tidak bisa mendeteksi keberadaan mereka.
Sekresi Sitokin Inhibitor: Melepaskan zat kimia yang membuat sel imun di sekitar tumor menjadi tidak aktif (anergik).
Tahapan:
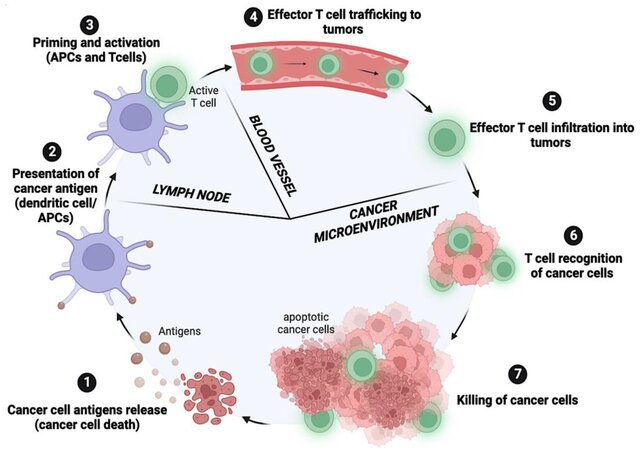
Ilustrasi skematis siklus kanker–imunitas, yang mewakili tujuh langkah kunci dalam mengembangkan respons imun terhadap kanker: (1) Antigen tumor dilepaskan; (2) sel dendritik menangkap dan menyajikan antigen tumor kepada sel T; (3) sel T diaktifkan dan dibedakan menjadi sel T efektor; (4) sel T efektor ditelusuri ke sel tumor; (5) sel T efektor menginfiltrasi jaringan tumor; (6) sel T mengenali sel kanker; (7) penekanan sel kanker.
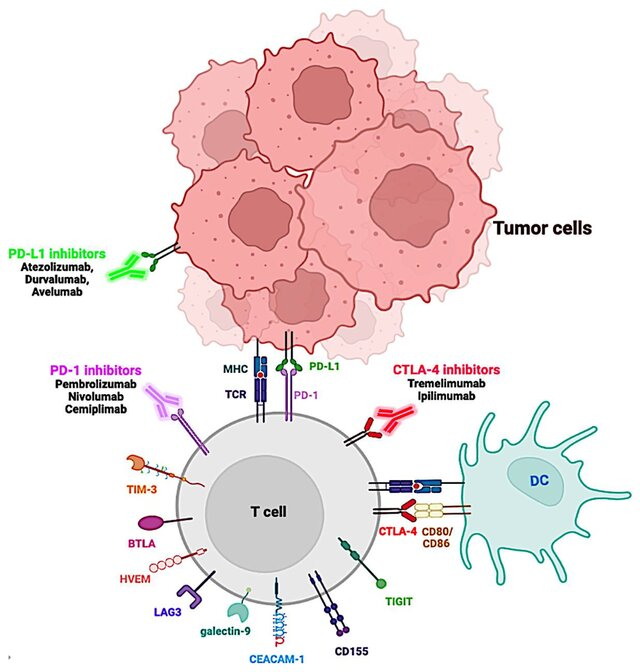
Inhibitor checkpoint imun yang disetujui FDA sebagai intervensi terapeutik dalam pengobatan kanker. Sel T diaktifkan melalui presentasi antigen oleh molekul kompleks histokompatibilitas utama (MHC) pada permukaan sel tumor atau pada sel penyaji antigen (APC) seperti sel dendritik (DC), yang dikenali oleh reseptor sel T (TCR). Sel T yang diaktifkan akan meningkatkan ekspresi reseptor permukaan sel ko-inhibitor seperti PD-1 dan CTLA-4. Pengikatan PD-1 ke ligan-ligannya, PD-L1, atau PD-L2, akan menghambat pensinyalan hilir TCR, sehingga menurunkan aktivitas sel T. Terapi antibodi yang menargetkan PD-1 atau PD-L1 dapat mengaktifkan kembali sel T yang kelelahan di lokasi tumor, meningkatkan aktivitasnya dan memungkinkan sel T untuk menghancurkan sel tumor. Pengikatan CTLA-4 ke ligan-ligannya, B7, CD80, atau CD86, menekan aktivitas sel T. Dengan memblokir interaksi CTLA-4 dengan antibodi anti-CTLA-4, proliferasi sel T akan diaktifkan.
Perbedaan Utama Kontrol Pembelahan
Fitur
Sel Imun (Normal)
Sel Kanker
Pemicu Belah
Antigen (Sinyal Eksternal)
Mutasi Onkogen (Sinyal Internal)
Siklus Sel
Berhenti setelah antigen hilang
Terus menerus (Abadi)
Stabilitas DNA
Mengatur DNA secara spesifik & presisi
Instabilitas genomik total
Apoptosis
Sensitif terhadap sinyal kematian
Resisten/Kebal terhadap apoptosis
KONTROL GENETIK RESPON IMUN
A. Komponen Sistem Imun Vertebrata
Ketika zat asing yang disebut antigen memasuki aliran darah mamalia, zat tersebut memicu mekanisme pertahanan, respons imun, yang menghasilkan sintesis kelompok protein yang sangat penting yang disebut antibodi. Antibodi ini mengikat antigen dengan spesifisitas yang luar biasa, sehingga memudahkan pembuangannya dari sistem peredaran darah. Selama beberapa dekade terakhir, para ilmuwan telah menemukan bahwa urutan DNA mengkodekan berbagai macam antibodi yang diproduksi oleh sistem imun mamalia disusun selama diferensiasi sel-sel penghasil antibodi melalui terjadinya serangkaian penataan ulang genom yang baru.
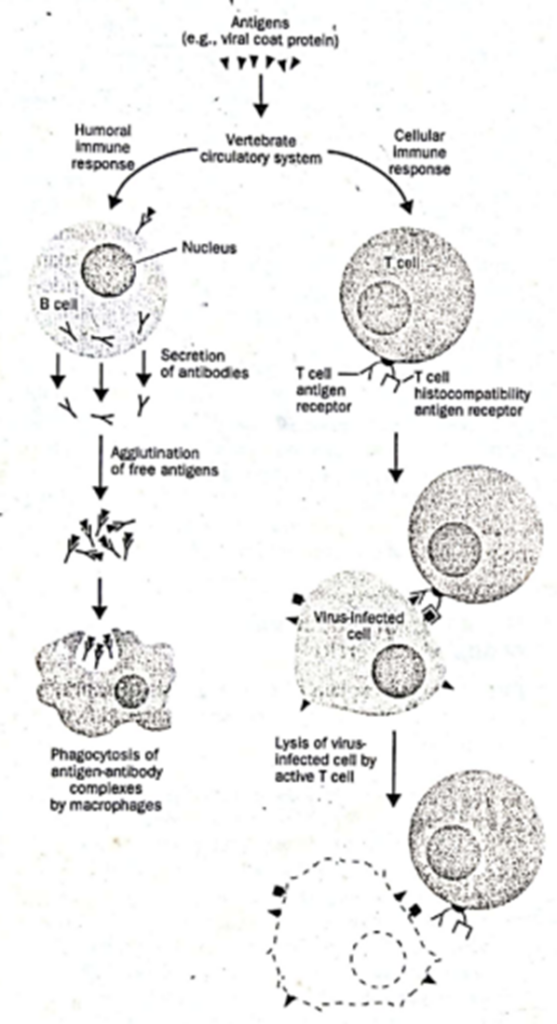
Adapun sistem imun didukung oleh beberapa komponen, seperti sel darah putih. Tiga jenis sel darah putih yang berbeda memainkan peran utama dalam respons imun pada vertebrata. Sel-sel ini adalah (1) limfosit B (disebut sel B karena diproduksi di sumsum tulang), (2) limfosit T (disebut sel T karena diproduksi di kelenjar timus), dan (3) makrofag. Antibodi disintesis oleh limfosit B dan disekresikan atau tetap terikat membran pada permukaan sel B tergantung pada kondisinya. Selama respons imun humoral, antibodi ini mengikat antigen bebas dalam sistem peredaran darah dan mengaglutinasinya. Kompleks antibodi-antigen yang dihasilkan kemudian dicerna dan didegradasi oleh makrofag (Gambar 1, kiri). Limfosit T memediasi respons imun seluler. Sel T mensintesis reseptor antigen yang mengenali antigen pada permukaan sel dan memicu lisis sel yang mengandung antigen oleh sel T yang diaktifkan (Gambar 1, kanan). Limfosit T yang berbeda melakukan fungsi ini dengan cara yang sedikit berbeda. Namun, secara umum, serangan sel T pada sel pembawa antigen memerlukan reseptor sel T spesifik dan satu atau lebih reseptor antigen histokompatibilitas.
Gambar 1. Diagram skematik komponen utama respons
Aspek yang paling luar biasa dari respons imun, setidaknya dari sudut pandang genetika, adalah respons yang tampaknya sebagai respons terhadap antigen pada hewan yang sebelumnya tidak ditemui. Bagaimana suatu organisme dapat mempersiapkan diri untuk mensintesis antibodi yang dirancang untuk mengikat secara sangat spesifik pada antigen tertentu tanpa pernah melakukan kontak dengan antigen tersebut.
B. Struktur Antibodi
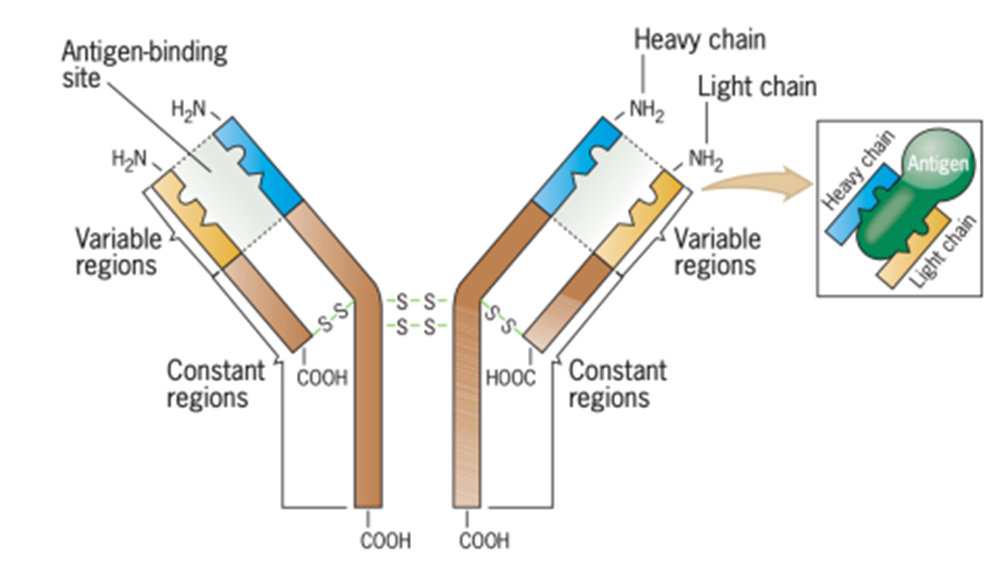
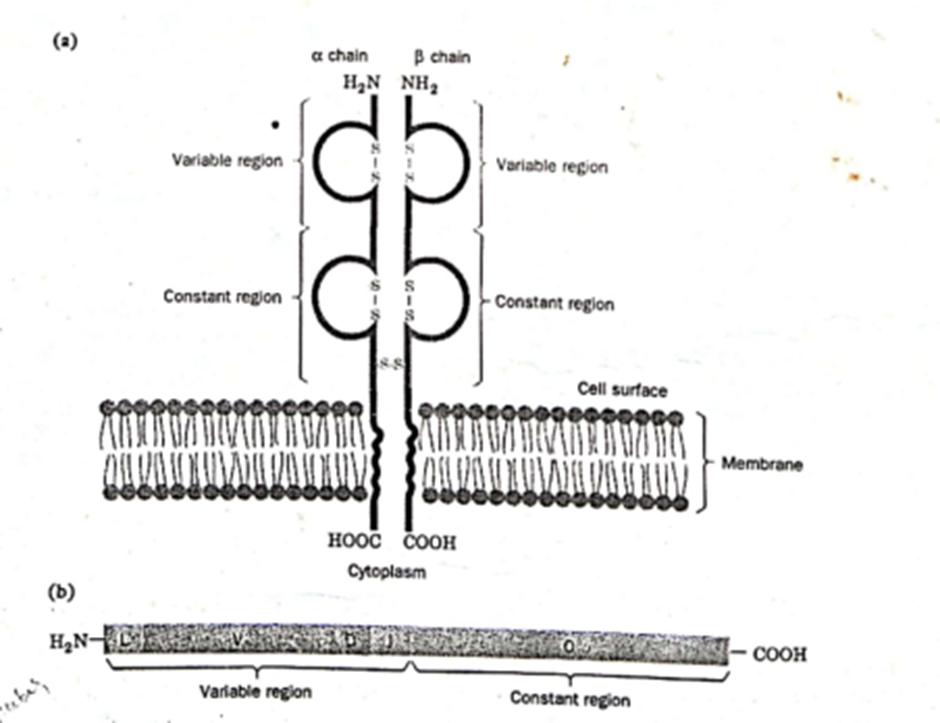
Antibodi termasuk dalam golongan protein yang disebut imunoglobulin. Setiap antibodi adalah tetramer yang terdiri dari empat polipeptida, dua light chains identik dan heavy chains identik, yang disatukan oleh ikatan disulfida (Gambar 2). Light chains panjangnya sekitar 220 asam amino, dan rantai berat panjangnya sekitar 440-450 asam amino. Setiap light and heavy chains memiliki amino-terminal variable region, di mana urutan asam amino bervariasi di antara antibodi yang spesifik untuk antigen yang berbeda, dan constant region karboksil terminal, di mana urutan asam amino sama untuk semua antibodi yang disebut kelas Imunoglobulin (Ig). Variable region dari semua rantai antibodi panjangnya sekitar 110 asam amino.
Gambar 2. Struktur molekul antibodi
(Sumber: Snustad)
Protein region yang menjalankan fungsi tertentu disebut domains. Setiap antibodi memiliki dua situs atau domains pengikat antigen, yang masing-masing dibentuk oleh daerah variabel dari satu light chains dan satu heavy chains. Selain itu, daerah konstan dari dua rantai berat berinteraksi untuk membentuk domain ketiga, yang disebut domain fungsi efektor, yang bertanggung jawab atas interaksi antibodi yang tepat dengan komponen lain dari sistem imun.
Ada lima kelas antibodi antara lain, IgM, IgD, IgG, IgE, dan IgA. Kelas tempat antibodi berada, dan dengan demikian fungsi yang dijalankannya, ditentukan oleh constant region pada
heavy chains (yaitu, struktur domain fungsi efektornya). Misalnya, antibodi IgD biasanya tetap terikat pada permukaan sel tempat mereka disintesis, sedangkan antitodi IgG biasanya disekresikan dan beredar melalui tubuh dalam aliran darah. Light chains antibodi terdiri dari dua jenis, kappa dan lambda, dengan jenis ditentukan oleh constant region pada light chains. Seperti yang akan kita lihat, antibodi mungkin memiliki spesifisitas pengikatan antigen yang sama, seperti yang ditentukan oleh daerah variabel dari keempat rantai, tetapi fungsi imunologis yang berbeda, seperti yang ditentukan oleh constant region pada heavy chains.
C. Hypothesis Antibody Diversity
Upaya-upaya terdahulu untuk menjelaskan dasar genetik keragaman antibodi dapat dikelompokkan secara garis besar ke dalam tiga hipotesis berbeda.
1. Hipotesis “germ line” menyatakan bahwa terdapat gen garis keturunan yang terpisah untuk setiap antibodi. Hal ini sesuai dengan pengetahuan awal tentang sintesis protein. 2. Hipotesis “somatic mutation” menyatakan bahwa hanya terdapat satu atau beberapa gen garis keturunan yang menentukan setiap kelas utama antibodi dan bahwa keragaman tersebut dihasilkan oleh frekuensi tinggi mutasi somatik.
3. Hipotesis “minigene” menyatakan bahwa keragaman tersebut dihasilkan oleh shuffling banyak segmen kecil dari beberapa gen menjadi banyak kemungkinan kombinasi. Shuffling terjadi melalui proses rekombinasi dalam sel somatik.
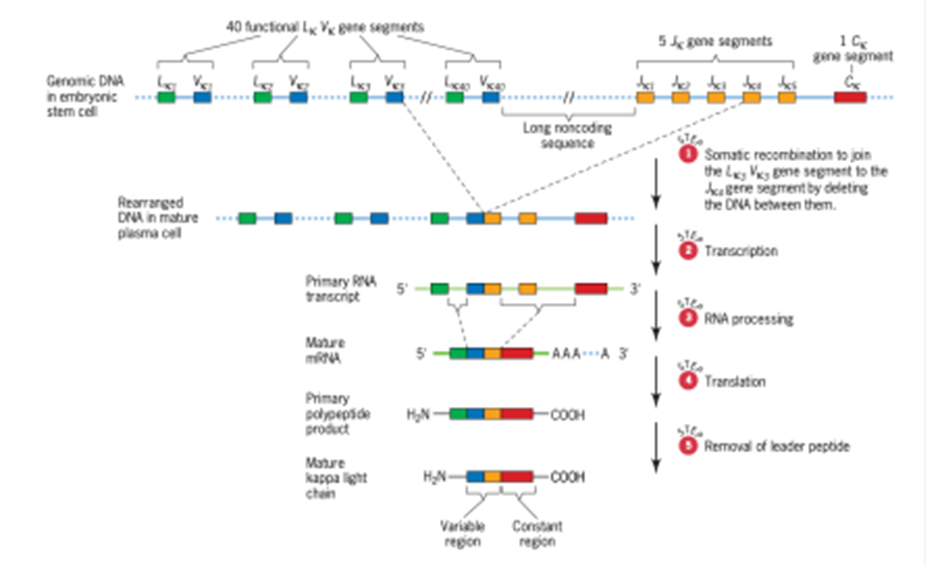
D. Diferensiasi Limfosit B (Light Chains, Heavy Chains, and Class Switching) The light and heavy chains dari antibodi dikodekan oleh lokus yang berbeda dalam genom. Pada manusia, terdapat dua lokus light chains, lokus kappa pada kromosom 2 dan lokus lambda pada kromosom 22, dan terdapat satu lokus heavy chains, yang terletak pada kromosom 14. Masing-masing lokus ini terdiri dari serangkaian segmen gen yang panjang. Lokus kappa menunjukkan bagaimana segmen-segmen ini diatur dan bagaimana mereka direkombinasi menjadi urutan pengkodean yang koheren untuk menghasilkan polipeptida yang berbeda (Gambar 3). Polipeptida kappa dikodekan oleh tiga jenis segmen gen:
1. Segmen gen LkVk, yang mengkode leader peptida dan 95 asam amino aminoterminal dari variable region kappa light chain; leader peptida dikeluarkan dari light chains kappa melalui pembelahan setelah memandu polipeptida yang baru terbentuk melalui membran retikulum endoplasma dalam sel plasma yang mensintesis antibodi.
2. Segmen gen Jk, yang mengkode 13 asam amino terakhir dari daerah variable region kappa light chain; simbol J digunakan untuk segmen gen ini karena peptida yang dikodenya
menggabungkan peptida terminal amino yang dikode oleh segmen LkVk ke peptida terminal karboksi yang dikode oleh jenis segmen gen berikutnya.
3. Segmen gen Ck, yang mengkode daerah konstan kappa light chain. Pada manusia, lokus kappa mengandung 76 segmen gen LkVk (meskipun hanya 40 yang fungsional), lima segmen gen Jk, dan satu segmen gen Ck. Segmen gen Jk terletak di antara segmen gen LkVk dan segmen gen Ck. Pada sel garis keturunan, lima segmen Jk dipisahkan dari segmen LkVk oleh urutan nonpengkodean yang panjang, dan dari segmen gen Ck oleh urutan nonpengkodean lain yang panjangnya sekitar 2 kb
Gambar 3. The genetic control of human antibody kappa light chains
(Sumber: Snustad)
E. Signal Sequence
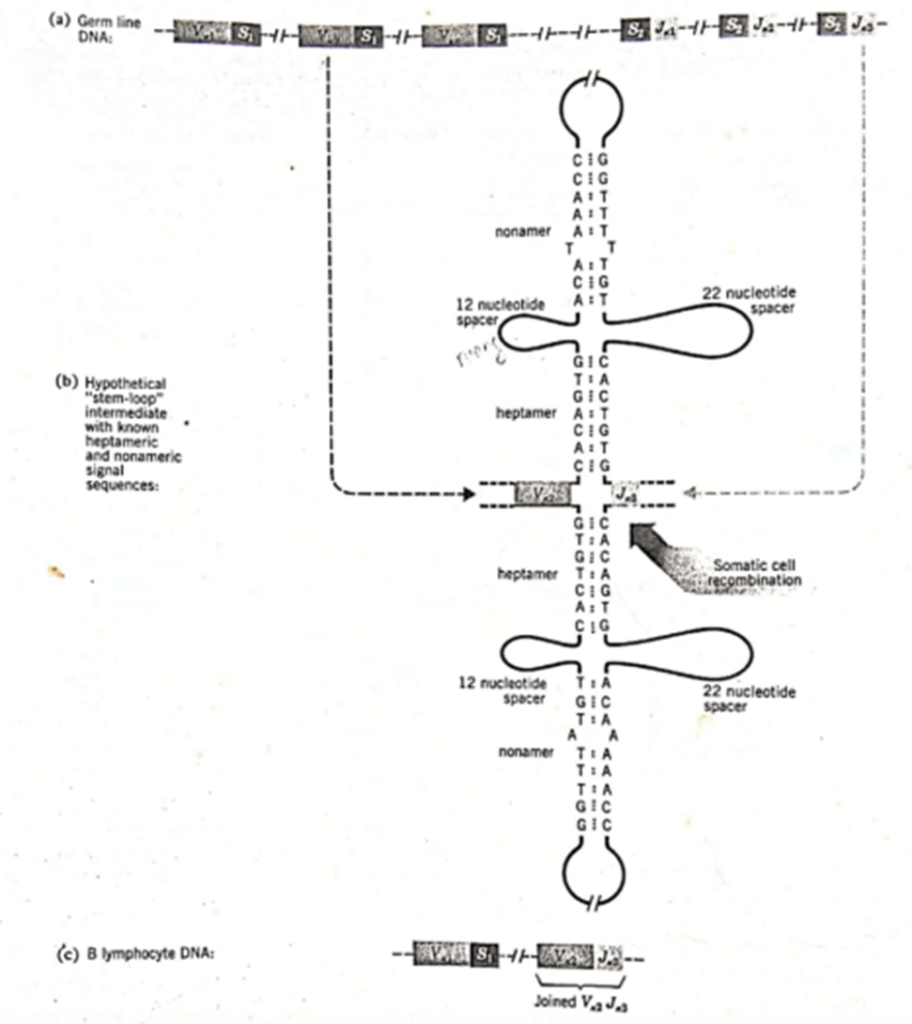
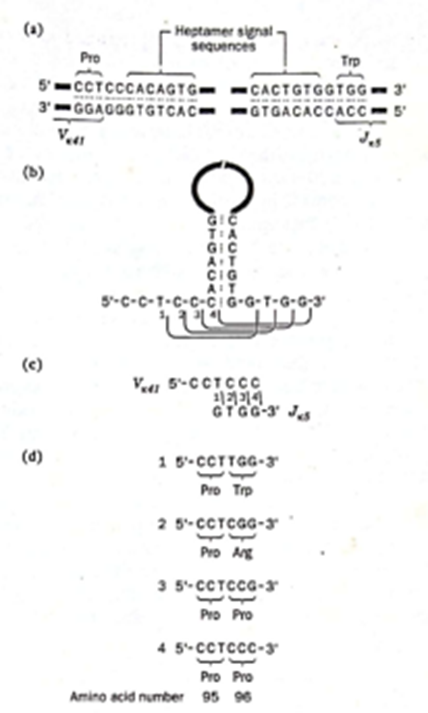
Bagaimana genome rearrangements yang terjadi selama perkembangan limfosit B diatur? Apa yang mengendalikan somatic recombination sehingga segmen gen V bergabung dengan segmen J dan bukan dengan segmen V lain atau langsung ke segmen C? Beberapa segmen panjang DNA kromosom yang membawa gugus segmen gen V, segmen gen D, dan segmen gen dari manusia kini telah diurutkan, dan urutan pasangan nukleotida yang dihasilkan menunjukkan adanya sinyal penggabungan V-J, V-D, dan D-J yang spesifik (Gambar 4). Urutan sinyal yang sama ditemukan berdekatan dengan semua segmen gen V. Demikian pula,
semua segmen gen J memiliki urutan sinyal identik yang terletak berdekatan dengan urutan pengkodeannya; namun, urutan sinyalnya berbeda dari yang berdekatan dengan segmen gen v. Demikian pula, segmen gen D dan C memiliki urutan sinyal yang berdekatan sendiri.
Gambar 4. Signal Sequence
(Sumber: Gardner)
F. Variable Joining and Somatic Mutations
Perbandingan keragaman urutan asam amino yang ada dalam molekul antibodi dengan yang diprediksi dari urutan segmen gen yang mengode antibodi ini mengungkapkan bahwa terdapat lebih banyak variasi dalam urutan asam amino pada V-J junctions daripada urutan nukleotida. Penelitian selanjutnya menunjukkan bahwa sebagian besar keragaman tambahan ini dapat dijelaskan oleh variasi pada lokasi rekombinasi yang tepat selama peristiwa V-J joining events. Contoh penggunaan lokasi penggabungan alternatif segmen gen Vk dan Jk pada diilustrasikan dalam Gambar 5. Selama penggabungan segmen gen Vk41 dan J5, rekombinasi telah terbukti terjadi antara empat posisi nukleotida yang berdekatan di lokasi sambungan. Peristiwa rekombinasi ini menghasilkan empat urutan nukleotida berbeda yang mengode tiga asam amino berbeda pada posisi 96 dalam lights chains kappa tikus. Lebih lanjut, karena asam amino 96 terjadi di wilayah rantai antibodi yang terlibat dalam pengikatan antigen, peristiwa V-J joining events alternatif jenis ini tidak diragukan lagi berkontribusi secara signifikan terhadap keragaman spesifisitas antibodi yang diamati pada vertebrata.
Gambar 5. Variable Joining
(Sumber: Gardner)
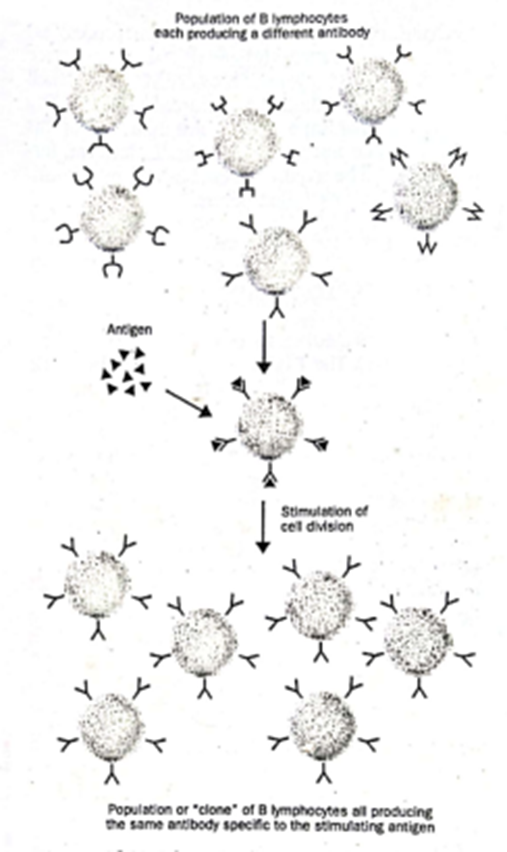
G. Clonal Selection
Seleksi klonal merupakan teori tentang bagaimana suatu organisme memulai sintesis antibodi yang spesifik terhadap antigen yang belum pernah ditemuinya sebelumnya. Semua antibodi yang diproduksi oleh satu limfosit B memiliki spesifitas pengikatan antigen yang sama. Namun, sel-sel yang berbeda dalam populasi limfosit B akan mengalami penataan ulang genom yang berbeda yang mengarah pada produksi antibodi dengan spesifitas yang berbeda. Dengan demikian, populasi limfosit B pada manusia atau tikus akan menghasilkan berbagai macam antibodi. Teori seleksi klonal menyatakan bahwa pengikatan antigen asing tertentu ke antibodi pada permukaan limfosit B merangsang sel tersebut untuk membelah, menghasilkan sejumlah besar limfosit B tertentu (“klon” dari sel-sel yang identik) dan dengan demikian sejumlah besar antibodi tertentu yang mengenali antigen asing (Gambar 6).
Gambar 6. Clonal Selection
(Sumber: Gardner)
H. T Cell Receptor
Limfosit T memediasi respons imun seluler. Sel T mengenali antigen pada permukaan sel dan membunuh sel yang membawa antigen ini. Seperti antibodi yang diproduksi oleh limfosit B, sel T dapat mengenali dan menghancurkan sel yang membawa berbagai macam antigen. Dengan demikian, respons sel T juga menunjukkan tingkat spesifisitas yang fenomenal. Sel T menghasilkan reseptor yang terikat membran yang sangat mirip dengan antibodi yang diproduksi oleh limfosit B. Selain itu, keragaman spesifisitas reseptor sel T dihasilkan oleh penataan ulang genom yang analog dengan yang terlibat dalam produksi antibodi.
Sel T harus secara bersamaan mengenali antigen yang menyerang pada permukaan sel dan protein lain yang hanya menempel pada permukaan sel. Protein permukaan sel kedua yang harus dikenali sel T ini adalah produk dari salah satu dari banyak gen dalam kompleks histocompatibility complex (MHC). Lokus MHC mengkodekan kelompok protein kompleks yang ada pada semua sel dalam tubuh manusia. Dengan demikian, sel T mampu mengenali dan menghancurkan sel apa pun yang memproduksi antigen tertentu di jaringan tubuh mana pun.
Reseptor sel T terdiri dari dua poli rantai peptida, α dan ß, masing-masing dikodekan oleh segmen gen L-V, D, J, dan C seperti rantai antibodi. Polipeptida α dan ß, seperti rantai antibodi, mengandung variable regions yang membentuk situs pengikatan antigen dan constant regions yang mengikat reseptor pada permukaan sel (Gambar 7). Variable regions reseptor sel T dikodekan oleh beberapa L-V, D, dan J, constant regions dikodekan oleh segmen gen C. Gen reseptor sel T disusun oleh genomic rearrangements yang terjadi selama diferensiasi limfosit T dari sel induk seperti halnya gen antibodi dalam limfosit B yang sedang berkembang. Protein reseptor α dan ß, dikodekan oleh segmen gen yang berjejer dalam kelompok, pada kromosom yang mirip dengan yang mengkode rantai antibodi. Pada manusia, kelompok segmen gen α dan ß, masing-masing terletak pada kromosom 14 dan 7. (Kelompok segmen gen lain mengkode jenis ketiga polipeptida reseptor sel T yang disebut γ yang terdapat pada jenis sel T tertentu. Ada beberapa jenis sel T berbeda yang menjalankan fungsi berbeda selama respons imun.)
Gambar 7. T Cell Receptor
(Sumber: Gardner)
KONTROL GENETIK PEMBELAHAN SEL KANKER
A. Regulasi Pembelahan Sel dan Terjadinya Kanker
Siklus sel terdiri dari periode pertumbuhan, sintesis DNA, dan pembelahan. Panjang siklus ini dan durasi masing-masing fasenya dikendalikan oleh sinyal kimia eksternal dan internal. Transisi dari setiap fase siklus memerlukan integrasi sinyal kimia tertentu dan respons yang tepat terhadap sinyal-sinyal ini. Jika sinyal-sinyal tersebut tidak dirasakan dengan benar atau jika sel tidak dipersiapkan dengan baik untuk merespons, sel tersebut dapat menjadi kanker.
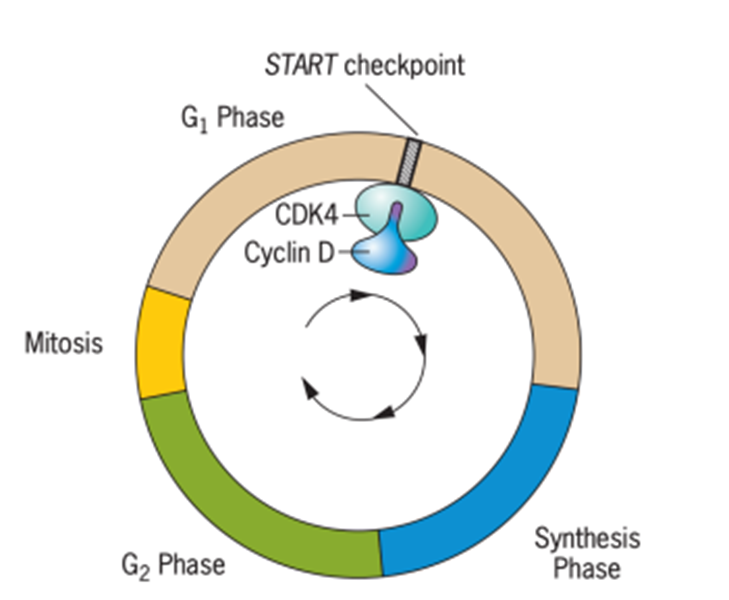
Pandangan terkini tentang kontrol siklus sel adalah bahwa transisi antara berbagai fase siklus (G1, S, G2, dan M diatur di “checkpoints”) (Gambar 8). Checkpoints adalah mekanisme yang menghentikan perkembangan melalui siklus hingga proses kritis seperti sintesis DNA selesai, atau hingga DNA yang rusak diperbaiki. Ketika checkpoints terpenuhi, siklus sel dapat berlanjut. Dua jenis protein memainkan peran penting dalam perkembangan ini, yaitu cyclins
and cyclin-dependent kinases, yang sering disingkat CDK. Kompleks yang terbentuk antara siklin dan CDK menyebabkan siklus sel berlanjut.
Gambar 8. Regulasi Pembelahan Sel
(Sumber: Snustad)
CDK merupakan komponen aktif katalitik dari mekanisme siklus sel. Protein ini mengatur aktivitas protein lain dengan mentransfer gugus fosfat kepada protein tersebut. Namun, aktivitas fosforilasi CDK bergantung pada keberadaan siklin. Siklin memungkinkan CDK menjalankan fungsinya dengan membentuk kompleks siklin/CDK. Jika siklin tidak ada, kompleks ini tidak dapat terbentuk, dan CDK tidak aktif. Oleh karena itu, siklus sel memerlukan pembentukan dan degradasi kompleks siklin/CDK secara bergantian.
Salah satu checkpoints siklus sel yang paling penting, disebut START, berada di pertengahan G1. Sel menerima sinyal eksternal dan internal di checkpoints ini untuk menentukan kapan waktu yang tepat untuk pindah ke fase S. Titik pemeriksaan ini diatur oleh siklin tipe D bersama dengan CDK4. Jika sel didorong melewati titik pemeriksaan START oleh kompleks siklin D/CDK4, sel tersebut berkomitmen pada putaran replikasi DNA lainnya. Protein penghambat dengan kemampuan merasakan masalah pada fase G1 akhir, seperti kadar nutrisi yang rendah atau kerusakan DNA, dapat mengerem kompleks siklin/CDK dan mencegah sel memasuki fase S. Jika tidak ada masalah seperti itu, kompleks siklin D/CDK4 mendorong sel melewati akhir fase G1 dan memasuki fase S, dengan demikian memulai replikasi DNA yang merupakan awal dari pembelahan sel.
Pada sel tumor, titik pemeriksaan dalam siklus sel biasanya mengalami deregulasi. Deregulasi ini disebabkan oleh cacat genetik dalam mesin yang secara bergantian menaikkan dan menurunkan kelimpahan kompleks siklin/CDK. Misalnya, gen yang mengkode siklin atau CDK dapat bermutasi, atau gen yang mengkode protein yang merespons kompleks siklin/CDK tertentu atau yang mengatur kelimpahan kompleks ini dapat bermutasi. Banyak jenis cacat genetik yang dapat menderegulasi siklus sel, dengan konsekuensi akhir bahwa sel dapat menjadi kanker.
Sel-sel yang checkpoints START-nya tidak berfungsi sangat rentan menjadi kanker. Checkpoints START mengendalikan masuknya ke fase S dari siklus sel. Jika DNA di dalam sel telah rusak, penting untuk menunda masuknya ke fase S agar DNA yang rusak dapat diperbaiki. Jika tidak, DNA yang rusak akan direplikasi dan ditransmisikan ke semua keturunan sel. Sel
sel normal diprogram untuk berhenti sejenak di checkpoints START untuk memastikan bahwa perbaikan selesai sebelum replikasi DNA dimulai. Sebaliknya, sel-sel yang titik pemeriksaan START-nya tidak berfungsi masuk ke fase S tanpa memperbaiki DNA yang rusak. Selama serangkaian siklus sel, mutasi yang diakibatkan oleh replikasi DNA yang tidak diperbaiki dapat terakumulasi dan menyebabkan deregulasi lebih lanjut dari siklus sel. Oleh karena itu, klon sel dengan titik pemeriksaan START yang tidak berfungsi dapat menjadi kanker yang agresif.
B. Peran Gen-Gen Tumor Suppressor dalam Kontrol Genetik Pembelahan Sel Hasil penelitian menunjukkan hanya sekitar 1 persen dari semua kanker bersifat turun temurun. Akan tetapi, lebih dari 20 sindrom kanker turunan yang berbeda telah diidentifikasi, dan pada hampir semuanya, cacat yang mendasarinya terdapat pada gen penekan tumor, bukan pada onkogen. Protein yang dikodekan oleh gen penekan tumor ini berfungsi dalam berbagai macam proses seluler, termasuk pembelahan, diferensiasi, kematian sel terprogram, dan perbaikan DNA. Pada bagian berikut, beberapa jenis protein tumor suppressor yang telah dipelajari secara intensif.
1) pRB Tumor Suppressor
Penelitian terkini telah mengungkapkan bahwa protein RB (pRB) tumor suppressor berperan penting dalam pengaturan siklus sel. Meskipun gen RB ditemukan melalui hubungannya dengan retinoblastoma, mutasi pada gen ini juga dikaitkan dengan jenis kanker lain, termasuk karsinoma paru sel kecil, osteosarkoma, serta karsinoma kandung kemih, serviks, dan prostat. Lebih jauh lagi, tikus yang homozigot untuk mutasi knockout RB mati selama perkembangan embrio. Dengan demikian, produk gen RB sangat penting bagi kehidupan.
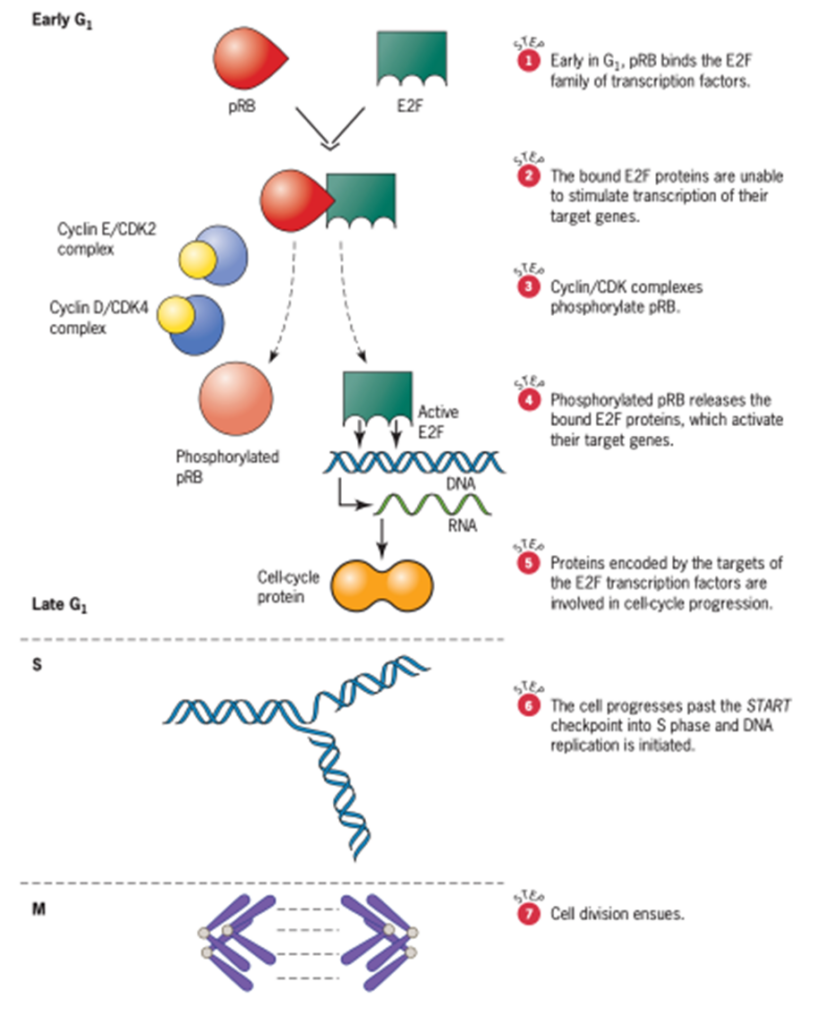
Analisis molekular dan biokimia telah menjelaskan peran pRB dalam regulasi siklus sel (Gambar 9). Pada awal fase G1 siklus sel, pRB mengikat protein E2F, faktor transkripsi yang mengendalikan ekspresi beberapa gen yang produknya menggerakkan sel melalui siklusnya. Ketika faktor transkripsi E2F terikat pada pRB, mereka tidak dapat mengikat urutan penambah spesifik dalam gen targetnya. Akibatnya, faktor siklus sel yang dikodekan oleh gen-gen ini tidak diproduksi, dan mesin untuk sintesis DNA dan pembelahan sel tetap diam. Kemudian pada G1, pRB difosforilasi melalui aksi kinase yang bergantung pada siklin. Dalam keadaan yang berubah ini, pRB melepaskan faktor transkripsi E2F yang telah terikat padanya. Faktor transkripsi yang dilepaskan ini kemudian bebas untuk mengaktifkan gen targetnya, yang mengkode protein yang menginduksi sel untuk maju melalui fase S dan menjadi mitosis. Setelah mitosis, pRB didefosforilasi, dan masing-masing sel anak memasuki fase diam dari siklus sel baru.
Gambar 9. Peran pRB Tumor Suppressor
(Sumber: Snustad)
2) p53 Tumor Suppressor
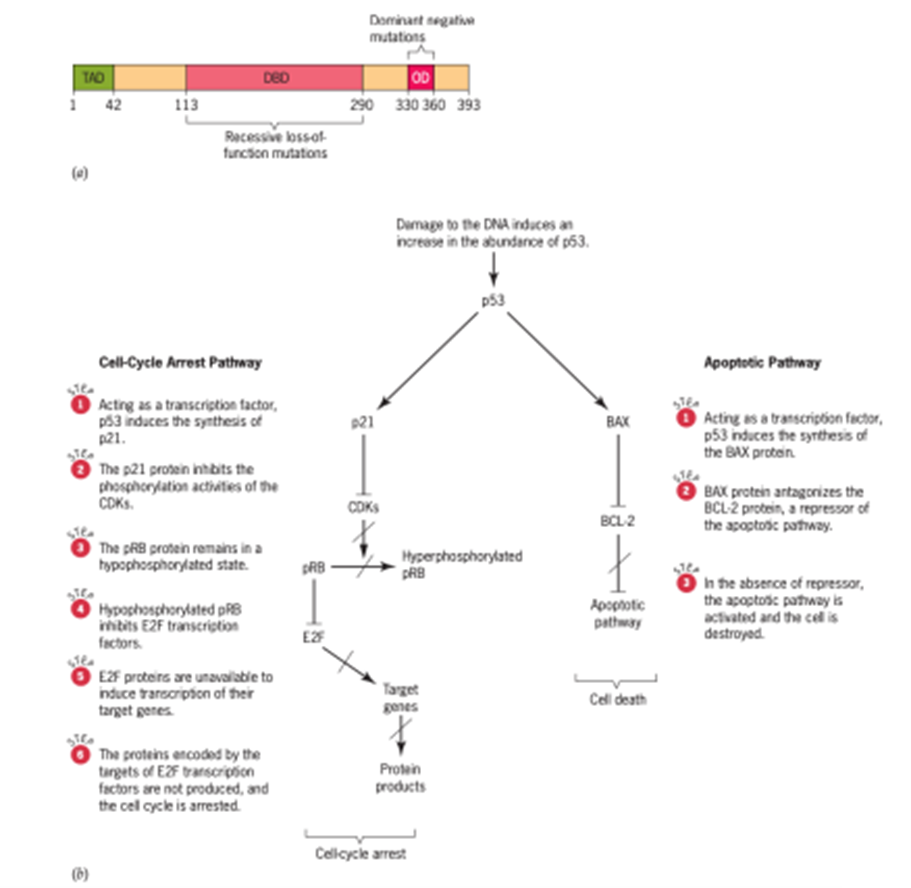
Protein p53 memainkan peran kunci dalam respons seluler terhadap stres (Gambar 10). Pada sel normal, kadar p53 rendah, tetapi ketika sel diobati dengan agen perusak DNA seperti radiasi, kadar p53 meningkat drastis. Respons terhadap kerusakan DNA ini dimediasi oleh jalur yang menurunkan degradasi p53. Sebagai respons terhadap kerusakan DNA, p53 difosforilasi, mengubahnya menjadi bentuk yang stabil dan aktif. Setelah diaktifkan, p53 menstimulasi transkripsi gen yang produknya menghentikan siklus sel, sehingga memungkinkan DNA yang rusak diperbaiki, atau mengaktifkan set gen lain yang produknya akhirnya menyebabkan sel yang rusak mati.
Gambar 10. p53 Tumor Suppressor
(Sumber: Snustad)
Protein p53 adalah faktor transkripsi sepanjang 393 asam amino yang terdiri dari tiga domain berbeda: N-terminal transcription-activation domain (TAD), DNA-binding core domain (DBD), dan C-terminal homo-oligomerization domain (OD). Sebagian besar mutasi yang menonaktifkan p53 terletak di DBD. Mutasi ini jelas merusak atau menghilangkan kemampuan p53 untuk mengikat urutan DNA spesifik yang tertanam dalam gen targetnya, sehingga mencegah aktivasi transkripsi gen-gen ini. Dengan demikian, mutasi pada DBD biasanya merupakan mutasi kehilangan fungsi resesif. Jenis mutasi lainnya ditemukan di bagian OD dari polipeptida. Molekul p53 dengan jenis mutasi ini dimerisasi dengan polipeptida p53 wild-type dan mencegah polipeptida wild-type berfungsi sebagai aktivator transkripsi. Dengan demikian, mutasi pada OD memiliki efek negatif dominan pada fungsi p53.
3) pAPC Tumor Suppressor
Protein pAPC memainkan peran kunci dalam mengatur pembaruan sel-sel pada lapisan, atau epitel, usus besar. Meskipun mekanisme yang mengatur proses ini tidak sepenuhnya dipahami, informasi terkini menunjukkan bahwa pAPC mengendalikan proliferasi dan diferensiasi sel-sel pada epitel usus. Ketika fungsi pAPC hilang, sel-sel yang menghasilkan proyeksi seperti jari pada epitel usus tetap dalam keadaan tidak berdiferensiasi. Ketika sel-sel ini terus membelah, mereka menghasilkan lebih banyak jenisnya sendiri, dan peningkatan jumlah sel yang dihasilkan menyebabkan banyak tumor jinak kecil terbentuk di epitel usus. Tumor ini disebut polip atau adenoma, dan kecenderungan untuk membentuknya diwariskan sebagai kondisi dominan autosomal langka yang disebut poliposis adenomatosa familial (FAP). Di negara-negara Barat, frekuensi populasinya sekitar 1 dalam 7000.
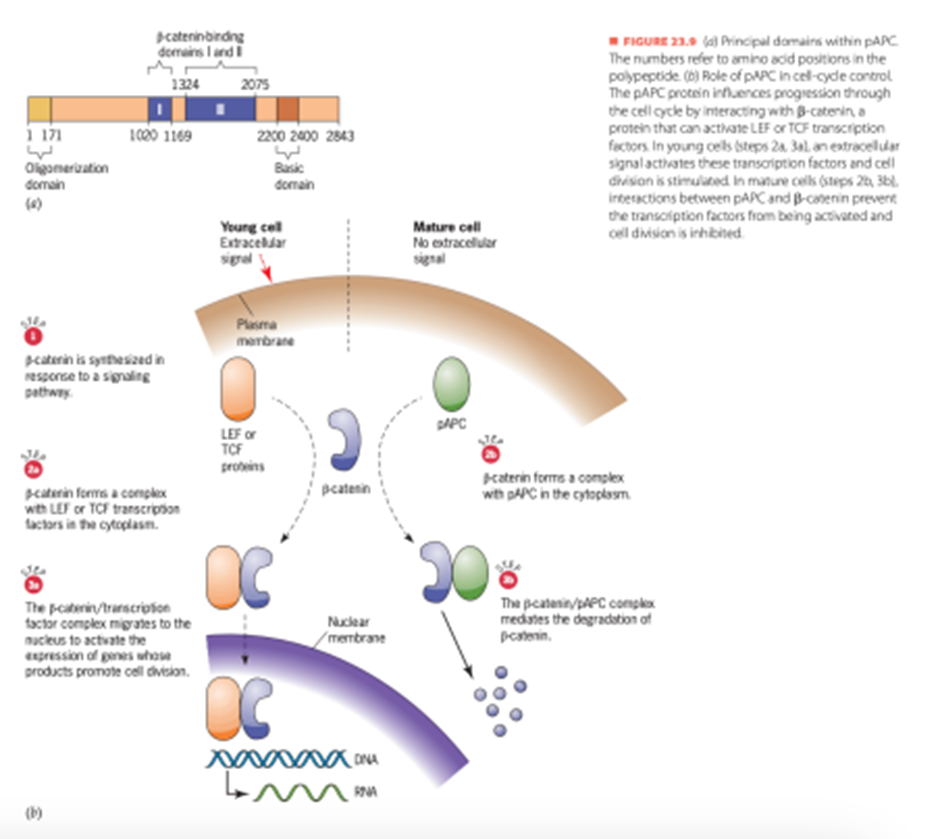
Selain itu, protein pAPC mengatur pembelahan sel melalui kemampuannya untuk mengikat β-catenin, protein yang ada di dalam sel. β-catenin secara alami juga mengikat protein lain, termasuk faktor transkripsi tertentu yang merangsang ekspresi gen yang produk proteinnya mendorong pembelahan sel. Interaksi dengan faktor transkripsi ini disukai ketika sinyal yang mengenai permukaan sel memberi isyarat kepada sel untuk membelah (Gambar 11). Proliferasi sel yang diinduksi sinyal merupakan proses yang diperlukan dalam epitel usus karena jaringan ini kehilangan sejumlah besar sel setiap hari pada manusia, sekitar 1011dan sel yang hilang harus digantikan oleh sel baru yang dihasilkan oleh pembelahan. Biasanya, sel-sel yang baru dibuat kehilangan kemampuan untuk membelah saat mereka menjauh dari bagian generatif epitel dan mengasumsikan peran mereka di bagian epitel yang matang. Pergeseran dari keadaan membelah menjadi tidak membelah terjadi karena sel-sel epitel yang matang tidak menerima sinyal ekstraseluler yang merangsang sel untuk membelah. Dengan tidak adanya
sinyal-sinyal ini, pAPC membentuk kompleks dengan β-catenin dalam sitoplasma sel, dan β catenin yang dikomplekskan menjadi sasaran degradasi. Karena pAPC menjaga kadar β catenin tetap rendah pada sel-sel epitel usus yang matang, ada sedikit peluang bagi β-catenin untuk bergabung dengan dan mengaktifkan faktor-faktor transkripsi yang merangsang pembelahan sel. Sel-sel dengan mutasi pada pAPC kehilangan kemampuannya untuk mengendalikan kadar β-catenin. Tanpa kendali ini, pAPC mempertahankan kekuatannya untuk membelah dan gagal berdiferensiasi dengan baik menjadi sel-sel epitel yang matang. Hasilnya adalah tumor mulai terbentuk di lapisan usus. Dengan demikian, molekul pAPC normal berperan penting dalam menekan pembentukan tumor di usus.
Gambar 11. pAPC Tumor Suppressor
(Sumber: Snustad)
C. Genetic Pathways to Cancer
Pada sebagian besar kasus kanker, pembentukan tumor ganas tidak disebabkan oleh aktivasi protoonkogen tunggal yang tidak terkendali atau inaktivasi gen penekan tumor tunggal. Sebaliknya, pembentukan, pertumbuhan, dan metastasis tumor biasanya bergantung pada akumulasi mutasi pada beberapa gen yang berbeda. Dengan demikian, jalur genetik menuju kanker beragam dan kompleks.
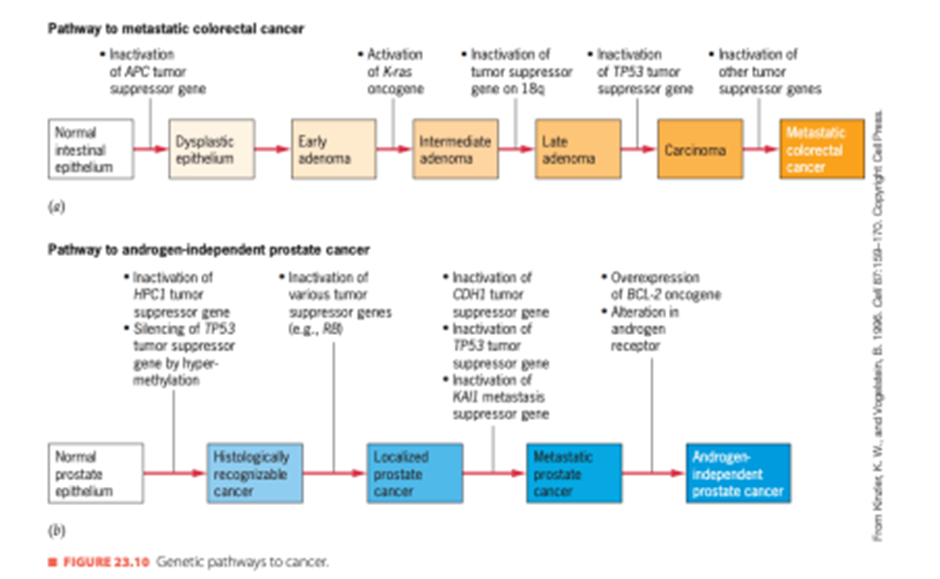
Keragaman dan kompleksitas ini terlihat dalam pembentukan dan perkembangan berbagai jenis tumor. Misalnya, tumor jinak usus besar berkembang pada individu dengan mutasi inaktivasi pada gen APC. Namun, perkembangan tumor ini menjadi kanker yang berpotensi mematikan memerlukan mutasi pada beberapa gen lainnya. Jalur mutasi ini dirangkum dalam Gambar 12a. Mutasi inaktivasi pada gen APC memulai proses pembentukan tumor dengan menyebabkan perkembangan jaringan abnormal di dalam epitel usus. Jaringan abnormal ini mengandung sel displastik, sel dengan bentuk yang tidak biasa dan inti yang membesar yang dapat tumbuh menjadi adenoma tahap awal. Jika proto-onkogen K-ras diaktifkan pada salah satu adenoma ini, adenoma tersebut dapat tumbuh dan berkembang lebih lengkap. Mutasi inaktivasi pada salah satu dari beberapa gen penekan tumor yang terletak di lengan panjang kromosom 18 kemudian dapat menyebabkan adenoma berkembang lebih lanjut, dan mutasi inaktivasi pada gen penekan tumor TP53 pada kromosom 17 dapat mengubahnya menjadi karsinoma yang tumbuh dengan cepat. Mutasi gen penekan tumor tambahan dapat memungkinkan sel karsinoma melepaskan diri dan menyerang jaringan lain. Dengan demikian, tidak kurang dari tujuh mutasi independen (dua mutasi inaktivasi pada gen APC, satu mutasi aktivasi pada gen K-ras, dua mutasi inaktivasi pada gen penekan tumor pada kromosom 18, dan dua mutasi inaktivasi pada gen TP53) diperlukan untuk perkembangan karsinoma usus, dan masih banyak lagi mutasi yang mungkin diperlukan untuk metastasis karsinoma tersebut ke bagian tubuh lainnya.
Jalur genetik menuju kanker prostat juga telah dijelaskan Gambar 12b. Mutasi pada HPC1, gen untuk kanker prostat herediter yang terletak di lengan panjang kromosom 1, telah terlibat dalam asal tumor prostat. Mutasi pada gen penekan tumor lain yang terletak di kromosom 13, 16, 17, dan 18 dapat mengubah tumor prostat menjadi kanker metastasis, dan ekspresi berlebihan gen protoonkogen BCL-2 dapat membuat kanker ini kebal terhadap terapi deprivasi androgen, teknik standar untuk pengobatan kanker prostat. Hormon steroid andro
gen diperlukan untuk proliferasi sel-sel di epitel prostat. Jika tidak ada androgen, sel-sel ini diprogram untuk mati. Namun, sel-sel tumor prostat dapat memperoleh kemampuan untuk bertahan hidup jika tidak ada androgen, karena kelebihan produk gen BCL-2 menekan jalur
kematian sel terprogram. Kanker prostat yang telah berkembang ke tahap androgen hampir selalu berakibat fatal.
Gambar 12. Genetic Pathway to Cancer
(Sumber: Snustad)
D. Protooncogene and Celular Oncogene
Protein yang dikode oleh onkogen virus mirip dengan protein seluler dengan fungsi pengaturan yang penting. Banyak dari protein seluler ini diidentifikasi dengan mengisolasi homolog seluler dari onkogen virus. Homolog seluler dari onkogen virus disebut proto oncogene, atau onkogen seluler normal, yang dilambangkan c-onc. Oleh karena itu, homolog seluler dari v-src adalah c-src. Urutan pengkodean kedua gen ini sangat mirip, hanya berbeda dalam 18 nukleotida; v-src mengkodekan protein dengan 526 asam amino, dan c-src mengkodekan protein dengan 533 asam amino. Dengan menggunakan gen v-onc sebagai probe, gen c-onc lainnya telah diisolasi dari banyak organisme yang berbeda, termasuk manusia. Sebagai aturan, onkogen seluler ini menunjukkan konservasi yang cukup besar dalam struktur. Drosophila, misalnya, membawa homolog yang sangat mirip dari onkogen seluler vertebrata c-abl, c-erbB, c-fps, c-raf, c-ras, dan c-myb. Kesamaan onkogen dari spesies yang berbeda sangat menunjukkan bahwa protein yang dikodekannya terlibat dalam fungsi seluler yang penting.
Produk-produk dari c-oncs memainkan peran kunci dalam mengatur aktivitas sel. Akibatnya, mutasi pada salah satu gen ini dapat mengganggu keseimbangan biokimia dalam sel dan menempatkannya pada jalur untuk menjadi kanker. Studi terhadap berbagai jenis kanker manusia telah menunjukkan bahwa onkogen sel yang bermutasi dikaitkan dengan perkembangan keadaan kanker. Bukti pertama yang menghubungkan kanker dengan mutan c
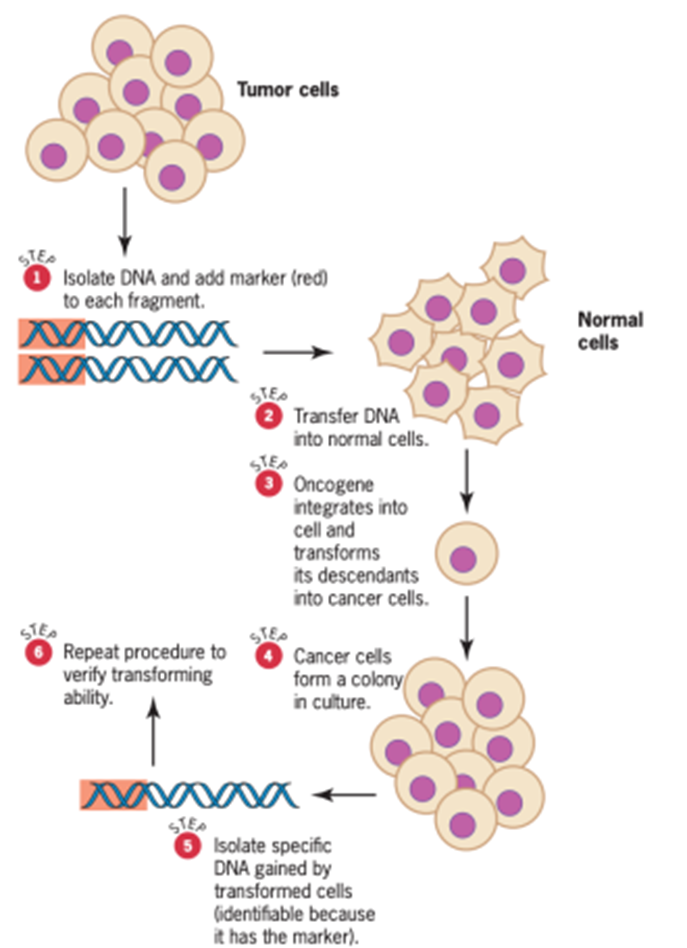
onc berasal dari penelitian kanker kandung kemih manusia. Mutasi yang menyebabkan kanker kandung kemih ini diisolasi oleh Robert Weinberg dan rekan-rekannya menggunakan uji transfeksi (Gambar 13).
Gambar 13. The transfection test to identify DNA sequences capable of transforming normal cells into cancer cells (Sumber: Snustad)
DNA diekstraksi dari jaringan kanker dan difragmentasi; kemudian setiap segmen disambungkan ke segmen DNA bakteri, yang berfungsi sebagai penanda molekuler. Fragmen DNA yang ditandai kemudian dimasukkan, atau ditransfeksi, ke dalam sel yang tumbuh dalam kultur untuk menentukan apakah ada segmen yang dapat mengubah sel menjadi keadaan kanker. Keadaan ini dapat dikenali dari kecenderungan sel kanker untuk membentuk gumpalan kecil, atau fokus, saat tumbuh soft agar plates. DNA dari sel-sel tersebut diekstraksi dan disaring untuk melihat apakah membawa penanda molekuler yang terkait dengan fragmen transfeksi asli. Jika ya, DNA ini diuji ulang untuk kemampuannya menginduksi keadaan kanker. Setelah beberapa pengujian, tim peneliti Weinberg mengidentifikasi fragmen DNA dari kanker kandung kemih asli yang secara reproduktif mengubah sel kultur menjadi sel kanker. Fragmen ini membawa alel onkogen c-H-ras, homolog onkogen dalam galur Harvey dari virus sarkoma tikus. Analisis urutan DNA kemudian menunjukkan bahwa nukleotida dalam kodon 12 dari alel ini telah bermutasi, dengan substitusi valin untuk glisin yang biasanya ditemukan pada posisi ini dalam protein c-H-ras.
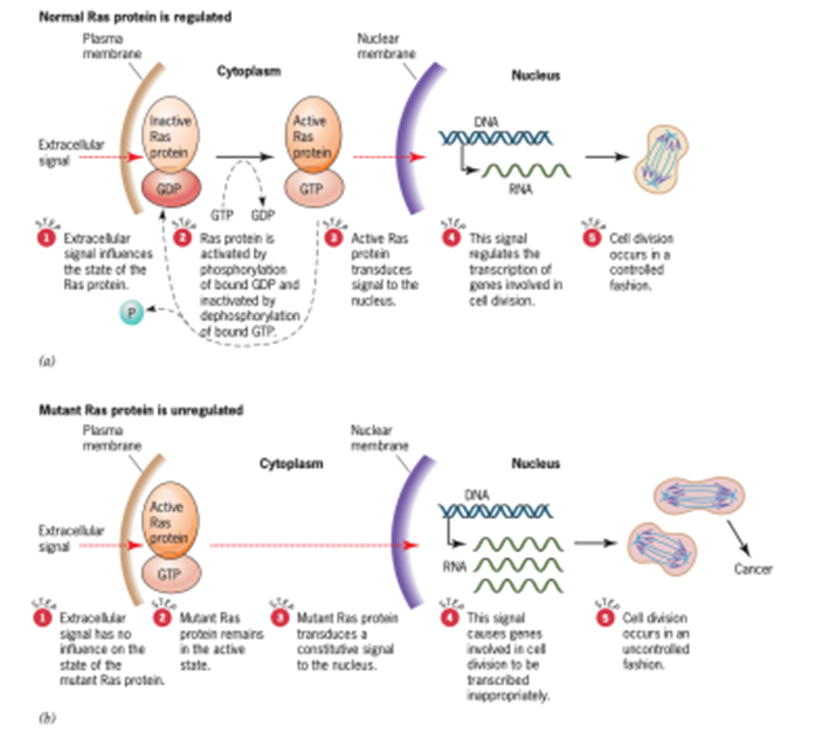
Ahli genetika kini memiliki pemahaman tentang bagaimana mutasi ini menyebabkan sel menjadi kanker. Tidak seperti onkogen virus, gen c-H-ras mutan tidak mensintesis protein dalam jumlah besar yang tidak normal. Sebaliknya, substitusi valin untuk glisin pada posisi 12 mengganggu kemampuan protein c-H-ras mutan untuk menghidrolisis salah satu substratnya, guanosine triphosphate (GTP). Lebih lanjut, karena gangguan ini, protein mutan tetap berada dalam mode pensinyalan aktif, mengirimkan informasi yang akhirnya merangsang sel untuk membelah dengan cara yang tidak terkendali (Gambar 14).
Gambar 14. Ras protein signaling and cancer
(Sumber: Snustad)
Versi mutan dari onkogen c-ras kini telah ditemukan dalam sejumlah besar tumor manusia yang berbeda, termasuk tumor paru-paru, usus besar, payudara, prostat, dan kandung kemih, serta neuroblastoma (kanker sel saraf), fibrosarkoma (kanker jaringan ikat), dan teratokarsinoma (kanker yang mengandung berbagai jenis sel embrionik). Dalam semua kasus, mutasi melibatkan perubahan asam amino dalam satu dari tiga posisi—12, 59, atau 61. Setiap perubahan asam amino ini merusak kemampuan protein Ras mutan untuk beralih dari mode pensinyalan aktifnya. Oleh karena itu, jenis mutasi ini merangsang sel untuk tumbuh dan membelah. Pada jenis kanker ini, hanya satu dari dua salinan gen c-ras yang telah bermutasi. Alel mutan tunggal dominan dalam kemampuannya untuk menimbulkan keadaan kanker. Mutasi pada c-ras dan onkogen seluler lainnya yang menyebabkan kanker dengan cara ini karenanya merupakan dominant activators pertumbuhan sel yang tidak terkendali.