Materi 1 – Regulasi Ekspresi Gen
- Pendahuluan Regulasi Ekspresi Gen
- Regulasi Ekspresi Gen Prokarotik
- Regulasi Ekspresi Gen Eukariot
- Video Pembelajaran
- Quiz
- Lembar Kerja Mahasiswa (LKM)
Pengertian Regulasi Ekspresi Gen
Ekspresi gen
Ekspresi gen mengacu pada proses pembuatan protein menggunakan instruksi dari gen. DNA seseorang mencakup banyak gen yang memilikiinstruksi untuk membuat protein. Selain itu, bagian-bagian tertentu dari DNA bukanlah bagian dari gen tetapi penting untuk memastikan genberfungsi dengan baik. Bagian-bagian DNA ini memberikan arahan tentang di mana dalam tubuh setiap protein harus dibuat, kapan harusdibuat, dan berapa banyak yang harus dibuat.
Pada umumnya, setiap sel dalam tubuh seseorang mengandung DNA dan gen yang sama persis, tetapi di dalam sel individu, beberapa gen aktif(“diaktifkan”) sementara yang lain tidak. Perbedaan dalam cara gen digunakan (diekspresikan) untuk membuat protein adalah alasan mengapabagian-bagian tubuh Anda terlihat dan berfungsi berbeda. Misalnya, ekspresi gen pada otot berbeda dengan ekspresi gen pada saraf.
Ekspresi gen dapat berubah seiring bertambahnya usia. Selain itu, perilaku Anda, seperti merokok atau berolahraga, atau paparan di lingkungansekitar dapat memengaruhi ekspresi gen.
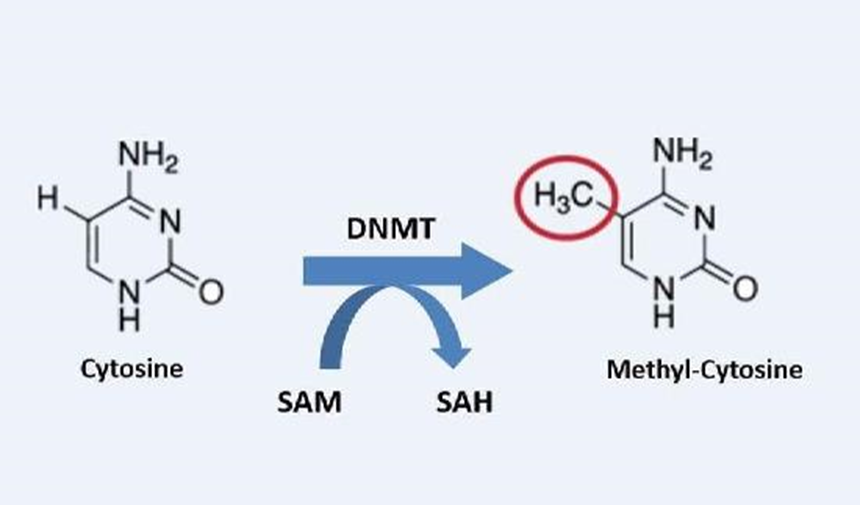
Gambar 1. Metilasi DNA
Metilasi DNA
Metilasi DNA bekerja dengan menambahkan zat kimia (yang dikenal sebagai gugus metil) ke DNA. Zat kimia ini juga dapat dihilangkan dari DNAmelalui proses yang disebut demetilasi. Biasanya, metilasi mematikan gen dan demetilasi mengaktifkan gen.
Metilasi DNA adalah salah satu cara tubuh mengontrol ekspresi gen. Metilasi dan demetilasi tidak mengubah kode DNA (urutan basa DNA),tetapi membantu menentukan seberapa banyak protein yang diproduksi.
Perubahan genetik (mutasi, varian gen, varian genetik)
Perubahan genetik (kadang-kadang disebut mutasi, varian gen, atau varian genetik) adalah perubahan dalam urutan basa DNA. Meskipun tidaksemua perubahan genetik akan menyebabkan masalah, terkadang, perubahan pada gen dapat menyebabkan perubahan pada protein dankemudian protein tersebut tidak berfungsi sebagaimana mestinya. Hal ini dapat menyebabkan penyakit.
Beberapa perubahan genetik dapat diturunkan dari orang tua kepada anak (diwariskan). Perubahan genetik ini terjadi pada sel germinal, yaitu selyang menghasilkan sperma atau sel telur. Perubahan genetik yang terjadi pada sel lain di dalam tubuh (dikenal sebagai sel somatik) tidakditurunkan kepada anak-anak seseorang.
Perubahan genetik terjadi ketika sel-sel baru dibuat dan DNA disalin. Selain itu, paparan, seperti radiasi tingkat tinggi, dapat merusak DNA danmenyebabkan perubahan genetik. Namun, sebagian besar paparan tidak akan mengakibatkan perubahan genetik karena setiap sel dalam tubuhmemiliki sistem untuk memeriksa kerusakan DNA dan memperbaiki kerusakan tersebut setelah ditemukan.
Regulasi Ekspresi gen Prokariotik dan Eukariotik
Regulasi ekspresi gen prokariotik berfokus pada efisiensi, terutama terjadi pada tingkat transkripsi untuk beradaptasi cepat dengan lingkungan. Mekanisme utamanya menggunakan sistem operon (induksi/represi) untuk mengatur kluster gen struktural melalui protein regulator (represor/aktivator) yang berikatan dengan operator atau promoter.
Berikut adalah poin-poin penting regulasi gen prokariotik:
- Sistem Operon: Unit fungsional yang terdiri dari gen struktural, promoter, dan operator (contoh: lac operon untuk metabolisme laktosa).
- Kontrol Negatif (Represor): Protein represor terikat pada operator, menghalangi RNA polimerase untuk mentranskripsi gen.
- Kontrol Positif (Aktivator): Protein aktivator membantu RNA polimerase berikatan dengan promoter, meningkatkan transkripsi.
- Induksi dan Represi: Gen dapat diinduksi (diaktifkan saat substrat ada) atau direpresi (dinonaktifkan saat produk akhir berlebih).
- Kopling Transkripsi-Translasi: Karena prokariot tidak memiliki inti sel, mRNA langsung ditranslasi saat transkripsi masih berjalan, sehingga regulasi pasca-transkripsi terbatas.
Regulasi ini memungkinkan bakteri seperti E. coli hanya memproduksi enzim yang diperlukan, menghemat energi secara efisien.
Regulasi ekspresi gen eukariotik adalah proses kompleks yang mengatur kapan, di mana, dan seberapa banyak gen diaktifkan. Regulasi terjadi di berbagai tahapan, terutama inisiasi transkripsi, melalui modifikasi kromatin, faktor transkripsi, splicing alternatif, dan kontrol pascatranslasi. Ini memastikan fungsi seluler yang tepat dan spesialisasi jaringan.
Berikut adalah tahapan regulasi ekspresi gen eukariotik:
- Tingkat Kromatin/Epigenetik (Pre-transkripsi): Mengatur aksesibilitas DNA. Kromatin yang padat (heterokromatin) tidak aktif, sementara yang longgar (eukromatin) aktif. Ini melibatkan asetilasi histon (membuka struktur) dan metilasi DNA (menutup/membungkam gen).
- Tingkat Transkripsi: Tahap utama regulasi. Faktor transkripsi (aktivator atau represor) berikatan dengan DNA pada elemen spesifik (enhancer atau promoter) untuk memacu atau menghambat RNA polimerase.
- Pemrosesan RNA (Pascatranskripsi): mRNA mentah (pre-mRNA) di dalam nukleus diproses. Alternative splicing memungkinkan satu gen menghasilkan protein yang berbeda dengan memotong intron dan menggabungkan ekson secara selektif.
- Stabilitas mRNA: Pengaturan berapa lama mRNA bertahan di sitoplasma sebelum didegradasi, mempengaruhi jumlah protein yang dihasilkan.
- Tingkat Translasi: Pengaturan kecepatan sintesis protein, seringkali melalui fosforilasi ribosom atau RNA interference (RNAi).
- Tingkat Pascatranslasi: Modifikasi protein setelah disintesis, seperti fosforilasi, glikosilasi, atau degradasi protein untuk mengaktifkan atau menonaktifkan fungsi protein tersebut.
Secara keseluruhan, regulasi eukariotik jauh lebih rumit daripada prokariotik karena pemisahan fisik transkripsi (di nukleus) dan translasi (di sitoplasma)
Regulasi Gen Prokariotik dan Eukariotik
Untuk memahami bagaimana ekspresi gen diatur, kita harus terlebih dahulu memahami bagaimana sebuah gen mengkode protein fungsional dalam sel. Proses ini terjadi baik pada sel prokariotik maupun eukariotik, hanya saja dengan cara yang sedikit berbeda.
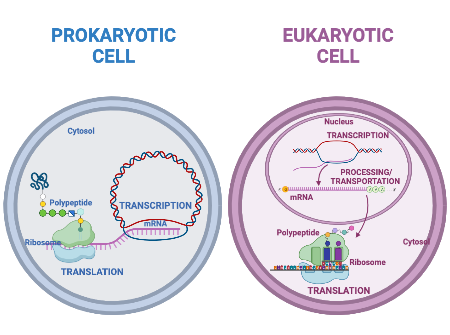
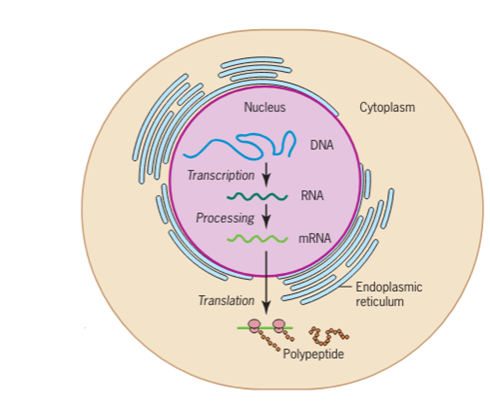
Organisme prokariotik adalah organisme bersel tunggal yang tidak memiliki inti sel, sehingga DNA mereka mengambang bebas di sitoplasma sel. Untuk mensintesis protein, proses transkripsi dan translasi terjadi hampir bersamaan. Ketika protein yang dihasilkan tidak lagi dibutuhkan, transkripsi berhenti. Akibatnya, metode utama untuk mengontrol jenis protein dan jumlah masing-masing protein yang diekspresikan dalam sel prokariotik adalah regulasi transkripsi DNA. Semua langkah selanjutnya terjadi secara otomatis. Ketika lebih banyak protein dibutuhkan, lebih banyak transkripsi terjadi. Oleh karena itu, dalam sel prokariotik, kontrol ekspresi gen sebagian besar berada pada tingkat transkripsi.
Sel eukariotik, sebaliknya, memiliki organel intraseluler yang menambah kompleksitasnya. Pada sel eukariotik, DNA terkandung di dalam nukleus sel dan di sana ditranskripsikan menjadi RNA. RNA yang baru disintesis kemudian diangkut keluar dari nukleus ke sitoplasma, tempat ribosom menerjemahkan RNA menjadi protein. Proses transkripsi dan translasi secara fisik dipisahkan oleh membran inti; transkripsi hanya terjadi di dalam nukleus, dan translasi hanya terjadi di luar nukleus di sitoplasma. Regulasi ekspresi gen dapat terjadi pada semua tahapan proses (Gambar 1). Regulasi dapat terjadi ketika DNA dilepaskan dan terlepas dari nukleosom untuk mengikat faktor transkripsi ( tingkat epigenetik ), ketika RNA ditranskripsikan (tingkat transkripsional), ketika RNA diproses dan diekspor ke sitoplasma setelah ditranskripsikan ( tingkat pasca-transkripsional ), ketika RNA diterjemahkan menjadi protein (tingkat translasional), atau setelah protein dibuat ( tingkat pasca-translasional ).
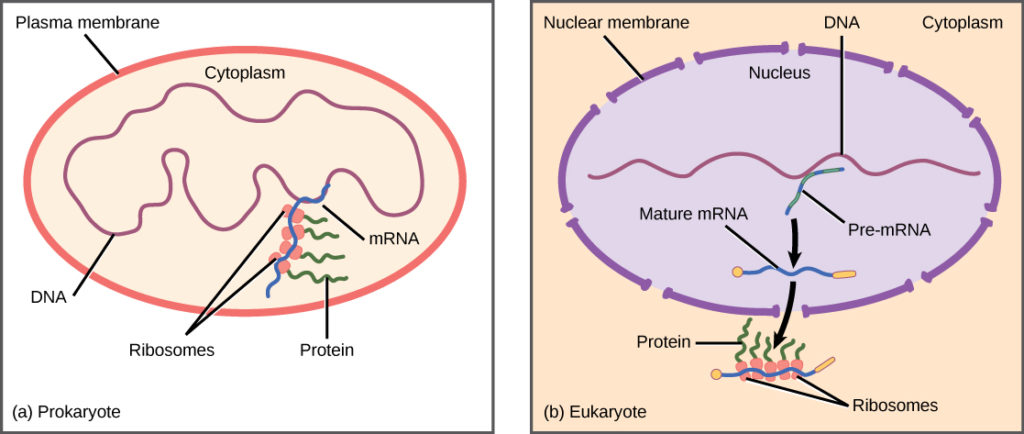
Gambar 1. Transkripsi dan translasi prokariotik terjadi secara bersamaan di sitoplasma, dan regulasi terjadi pada tingkat transkripsi. Ekspresi gen eukariotik diatur selama transkripsi dan pemrosesan RNA, yang terjadi di nukleus, dan selama translasi protein, yang terjadi di sitoplasma. Regulasi lebih lanjut dapat terjadi melalui modifikasi pasca-translasi protein.
Perbedaan dalam regulasi ekspresi gen antara prokariota dan eukariota dirangkum dalam Tabel 1. Regulasi ekspresi gen dibahas secara rinci dalam modul-modul selanjutnya.

Regulasi Gen Prokariotik
DNA prokariota tersusun dalam kromosom melingkar yang tergulung rapat di wilayah nukleoid sitoplasma sel. Protein yang dibutuhkan untuk fungsi spesifik dikodekan bersama dalam blok yang disebut operon . Misalnya, semua gen yang dibutuhkan untuk menggunakan laktosa sebagai sumber energi dikodekan berdampingan dalam operon laktosa (atau lac).
Pada sel prokariotik, terdapat tiga jenis molekul pengatur yang dapat memengaruhi ekspresi operon: represor, aktivator, dan induser.
- Represor adalah protein yang menekan transkripsi suatu gen sebagai respons terhadap rangsangan eksternal, sedangkan
- aktivator adalah protein yang meningkatkan transkripsi suatu gen sebagai respons terhadap rangsangan eksternal. Terakhir,
- induser adalah molekul kecil yang mengaktifkan atau menekan transkripsi tergantung pada kebutuhan sel dan ketersediaan substrat.
Regulasi Gen pada Prokariota
Pada bakteri dan archaea, protein struktural dengan fungsi terkait—seperti gen yang mengkode enzim yang mengkatalisis banyak langkah dalam satu jalur biokimia—biasanya dikodekan bersama dalam genom dalam blok yang disebut operon dan ditranskripsikan bersama di bawah kendali satu promotor. Ini membentuk transkrip polikistronik (Gambar 1). Promotor kemudian memiliki kendali simultan atas pengaturan transkripsi gen struktural ini karena semuanya akan dibutuhkan pada waktu yang sama, atau tidak satupun yang dibutuhkan.
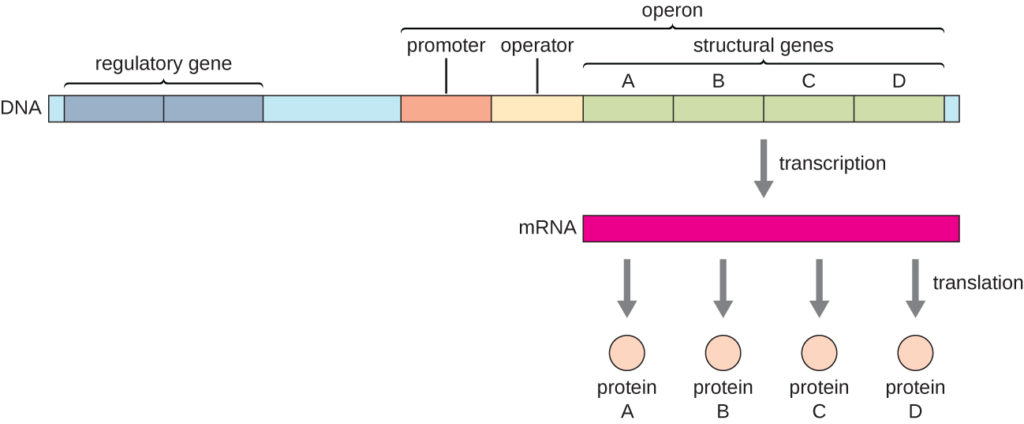
Gambar 1. Pada prokariota, gen struktural dengan fungsi terkait sering kali tersusun bersama pada genom dan ditranskripsikan bersama di bawah kendali satu promotor. Daerah pengatur operon mencakup promotor dan operator. Jika represor berikatan dengan operator, maka gen struktural tidak akan ditranskripsikan. Sebaliknya, aktivator dapat berikatan dengan daerah pengatur, sehingga meningkatkan transkripsi.
Ilmuwan Prancis François Jacob (1920–2013) dan Jacques Monod di Institut Pasteur adalah orang pertama yang menunjukkan pengorganisasian gen bakteri ke dalam operon, melalui studi mereka tentang operon lac pada E. coli . Mereka menemukan bahwa pada E. coli , semua gen struktural yang mengkode enzim yang dibutuhkan untuk menggunakan laktosa sebagai sumber energi terletak bersebelahan dalam operon laktosa (atau lac ) di bawah kendali satu promotor tunggal, yaitu promotor lac . Atas karya ini, mereka memenangkan Hadiah Nobel dalam Fisiologi atau Kedokteran pada tahun 1965.
Meskipun gen eukariotik tidak tersusun dalam operon, operon prokariotik merupakan model yang sangat baik untuk mempelajari regulasi gen secara umum. Terdapat beberapa gugus gen pada eukariota yang berfungsi mirip dengan operon. Banyak prinsip yang dapat diterapkan pada sistem eukariotik dan berkontribusi pada pemahaman kita tentang perubahan ekspresi gen pada eukariota yang dapat mengakibatkan perubahan patologis seperti kanker.
Setiap operon mencakup sekuens DNA yang memengaruhi transkripsinya sendiri; sekuens ini terletak di wilayah yang disebut wilayah pengatur. Wilayah pengatur mencakup promotor dan wilayah di sekitarnya, tempat faktor transkripsi , protein yang dikodekan oleh gen pengatur, dapat berikatan. Faktor transkripsi memengaruhi pengikatan RNA polimerase ke promotor dan memungkinkan progresinya untuk mentranskripsikan gen struktural. Represor adalah faktor transkripsi yang menekan transkripsi suatu gen sebagai respons terhadap stimulus eksternal dengan berikatan dengan sekuens DNA di dalam wilayah pengatur yang disebut operator , yang terletak di antara situs pengikatan RNA polimerase pada promotor dan situs awal transkripsi gen struktural pertama. Pengikatan represor secara fisik menghalangi RNA polimerase untuk mentranskripsikan gen struktural. Sebaliknya, aktivator adalah faktor transkripsi yang meningkatkan transkripsi suatu gen sebagai respons terhadap stimulus eksternal dengan memfasilitasi pengikatan RNA polimerase ke promotor. Induser , jenis molekul pengatur ketiga, adalah molekul kecil yang mengaktifkan atau menekan transkripsi dengan berinteraksi dengan represor atau aktivator.
Pada prokariota, terdapat contoh operon yang produk gennya dibutuhkan secara konsisten dan oleh karena itu ekspresinya tidak diatur. Operon semacam itu diekspresikan secara konstitutif , artinya ditranskripsikan dan diterjemahkan secara terus menerus untuk menyediakan sel dengan tingkat perantara produk protein yang konstan. Gen-gen tersebut mengkodekan enzim yang terlibat dalam fungsi pemeliharaan sel yang diperlukan untuk pemeliharaan seluler, termasuk replikasi, perbaikan, dan ekspresi DNA, serta enzim yang terlibat dalam metabolisme inti. Sebaliknya, ada operon prokariotik lain yang hanya diekspresikan ketika dibutuhkan dan diatur oleh represor, aktivator, dan induser.
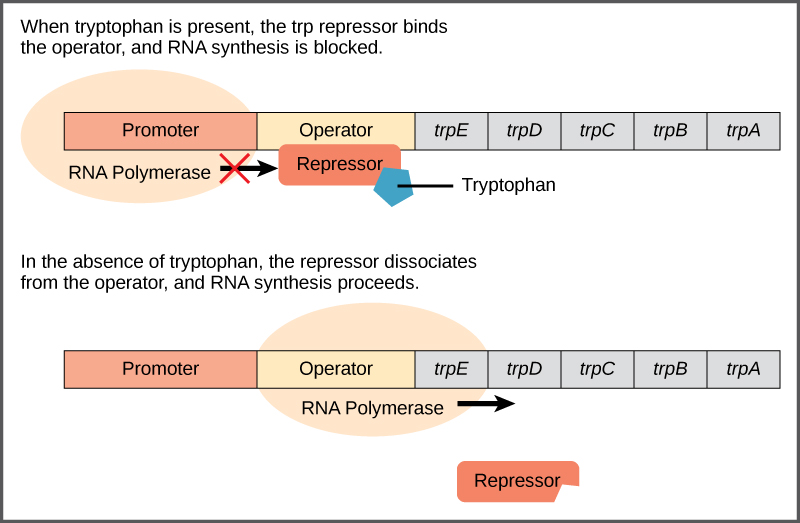
Operon trp : Sebuah Operon Represor
Bakteri seperti E. coli membutuhkan asam amino untuk bertahan hidup. Triptofan adalah salah satu asam amino yang dapat diserap E. coli dari lingkungan. E. coli juga dapat mensintesis triptofan menggunakan enzim yang dikodekan oleh lima gen. Kelima gen ini bersebelahan dalam apa yang disebut operon triptofan ( trp ) (Gambar 2). Jika triptofan ada di lingkungan, maka E. coli tidak perlu mensintesisnya dan saklar yang mengontrol aktivasi gen dalam operon trp dimatikan. Namun, ketika ketersediaan triptofan rendah, saklar yang mengontrol operon diaktifkan, transkripsi dimulai, gen diekspresikan, dan triptofan disintesis.
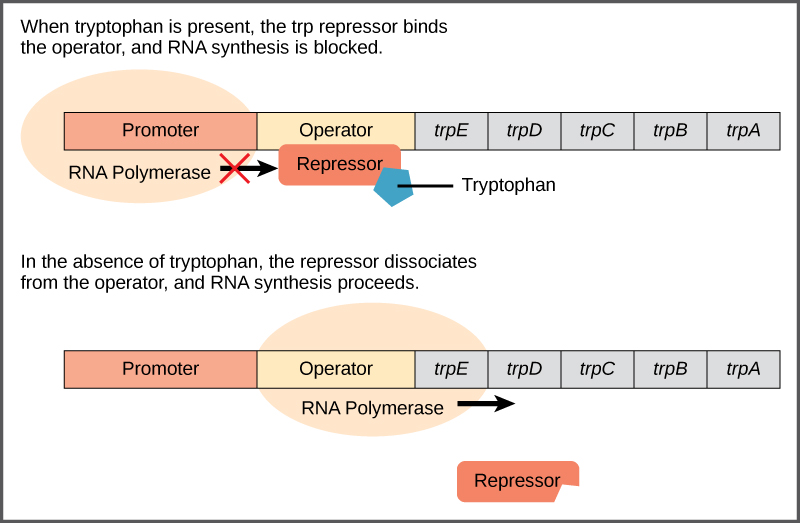
Gambar 2. Lima gen yang dibutuhkan untuk mensintesis triptofan pada E. coli terletak bersebelahan dalam operon
trp . Ketika triptofan melimpah, dua molekul triptofan mengikat protein represor pada sekuens operator. Hal ini secara fisik menghalangi RNA polimerase untuk mentranskripsikan gen triptofan. Ketika triptofan tidak ada, protein represor tidak mengikat operator dan gen tersebut ditranskripsikan.
Urutan DNA yang mengkode protein disebut sebagai daerah pengkodean. Lima daerah pengkodean untuk enzim biosintesis triptofan tersusun secara berurutan pada kromosom dalam operon. Tepat sebelum daerah pengkodean terdapat situs awal transkripsi . Ini adalah daerah DNA tempat RNA polimerase berikatan untuk memulai transkripsi. Urutan promotor berada di hulu situs awal transkripsi; setiap operon memiliki urutan di dalam atau di dekat promotor tempat protein (aktivator atau represor) dapat berikatan dan mengatur transkripsi.
Urutan DNA yang disebut urutan operator dikodekan di antara wilayah promotor dan gen pengkode trp pertama. Operator ini mengandung kode DNA yang dapat diikat oleh protein represor. Ketika triptofan hadir dalam sel, dua molekul triptofan mengikat represor trp , yang mengubah bentuknya untuk mengikat operator trp . Pengikatan kompleks triptofan-represor pada operator secara fisik mencegah RNA polimerase untuk mengikat dan mentranskripsikan gen-gen selanjutnya.
Ketika triptofan tidak ada di dalam sel, represor itu sendiri tidak mengikat operator; oleh karena itu, operon aktif dan triptofan disintesis. Karena protein represor secara aktif mengikat operator untuk menjaga agar gen tetap mati, operon trp diatur secara negatif dan protein yang mengikat operator untuk membungkam ekspresi trp adalah regulator negatif .
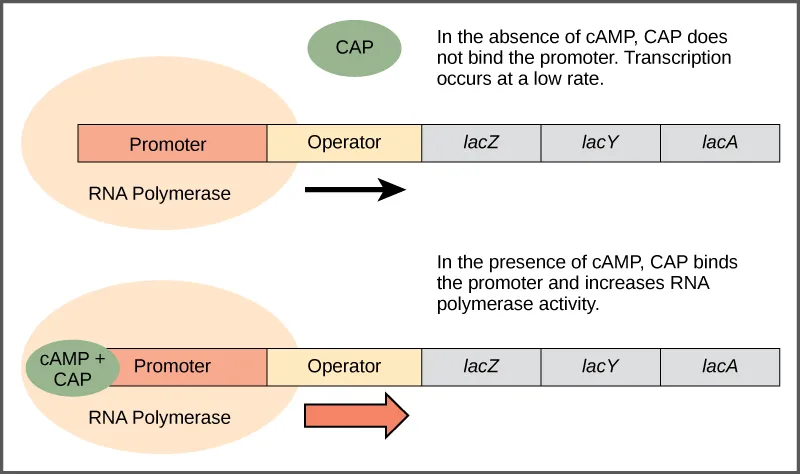
Protein Pengaktif Katabolit (CAP): Regulator Pengaktif
Sama seperti operon trp yang diatur secara negatif oleh molekul triptofan, terdapat protein yang mengikat sekuens operator yang bertindak sebagai regulator positif untuk mengaktifkan gen. Misalnya, ketika glukosa langka, bakteri E. coli dapat beralih ke sumber gula lain sebagai bahan bakar. Untuk melakukan ini, gen baru untuk memproses gen alternatif ini harus ditranskripsikan. Ketika kadar glukosa turun, AMP siklik (cAMP) mulai menumpuk di dalam sel. Molekul cAMP adalah molekul pensinyalan yang terlibat dalam metabolisme glukosa dan energi di E. coli . Ketika kadar glukosa menurun di dalam sel, cAMP yang menumpuk mengikat regulator positif protein aktivator katabolit (CAP) , protein yang mengikat promotor operon yang mengontrol pemrosesan gula alternatif. Ketika cAMP mengikat CAP, kompleks tersebut mengikat daerah promotor gen yang dibutuhkan untuk menggunakan sumber gula alternatif (Gambar 1). Dalam operon ini, situs pengikatan CAP terletak di hulu situs pengikatan RNA polimerase di promotor. Hal ini meningkatkan kemampuan pengikatan RNA polimerase ke wilayah promotor dan transkripsi gen.

Gambar 3. Ketika kadar glukosa menurun, E. coli dapat menggunakan gula lain sebagai bahan bakar tetapi harus mentranskripsikan gen baru untuk melakukannya. Saat pasokan glukosa terbatas, kadar cAMP meningkat. cAMP ini berikatan dengan protein CAP, regulator positif yang berikatan dengan daerah operator di hulu gen yang diperlukan untuk menggunakan sumber gula lain.
Operon lac : Sebuah Operon Induser
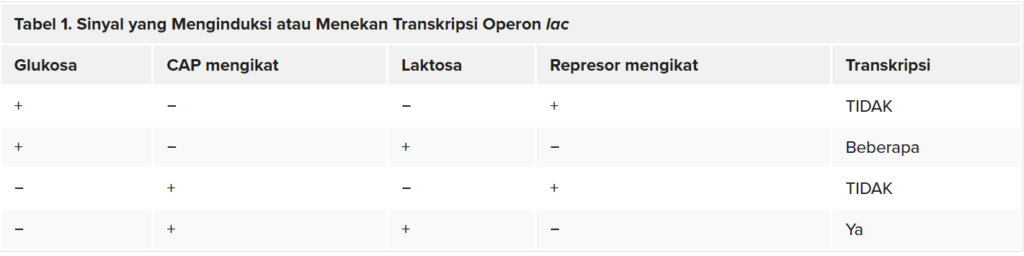
Jenis regulasi gen ketiga pada sel prokariotik terjadi melalui operon yang dapat diinduksi , yang memiliki protein yang mengikat untuk mengaktifkan atau menekan transkripsi tergantung pada lingkungan lokal dan kebutuhan sel. Operon lac adalah contoh operon yang dapat diinduksi. Seperti yang telah disebutkan sebelumnya, E. coli mampu menggunakan gula lain sebagai sumber energi ketika konsentrasi glukosa rendah. Untuk melakukan hal tersebut, kompleks protein cAMP–CAP berfungsi sebagai regulator positif untuk menginduksi transkripsi. Salah satu sumber gula tersebut adalah laktosa. Operon lac mengkode gen yang diperlukan untuk memperoleh dan memproses laktosa dari lingkungan lokal. CAP mengikat sekuens operator di hulu promotor yang memulai transkripsi operon lac . Namun, agar operon lac diaktifkan, dua kondisi harus dipenuhi. Pertama, kadar glukosa harus sangat rendah atau tidak ada. Kedua, laktosa harus ada. Hanya ketika glukosa tidak ada dan laktosa ada, operon lac akan ditranskripsikan. Hal ini masuk akal bagi sel, karena akan boros energi untuk membuat protein guna memproses laktosa jika glukosa berlimpah atau laktosa tidak tersedia.
Jika glukosa tidak ada, maka CAP dapat berikatan dengan sekuens operator untuk mengaktifkan transkripsi. Jika laktosa tidak ada, maka represor berikatan dengan operator untuk mencegah transkripsi. Jika salah satu dari persyaratan ini terpenuhi, maka transkripsi tetap nonaktif. Hanya ketika kedua kondisi terpenuhi, operon
lac ditranskripsikan (Tabel 1).
Regulasi Gen Prokariotik yang Sedang Beraksi
Seperti yang baru saja kita pelajari, ada tiga jenis molekul pengatur yang dapat memengaruhi ekspresi operon: represor, aktivator, dan inducer.
- Represor adalah protein yang menekan transkripsi suatu gen sebagai respons terhadap rangsangan eksternal. Dengan kata lain, represor menjaga agar gen tetap “mati”.
- Aktivator adalah protein yang meningkatkan transkripsi suatu gen sebagai respons terhadap rangsangan eksternal. Dengan kata lain, aktivator mengaktifkan suatu gen.
- Induser adalah molekul kecil yang mengaktifkan atau menekan transkripsi tergantung pada kebutuhan sel dan ketersediaan substrat. Pada dasarnya, induser membantu mempercepat atau memperlambat proses “aktif” atau “nonaktif” dengan berikatan dengan represor atau aktivator. Dengan kata lain: mereka tidak bekerja sendiri.
Regulasi Gen pada Eukariot
Genom manusia mengkode lebih dari 20.000 gen; setiap dari 23 pasang kromosom manusia mengkode ribuan gen. DNA di dalam nukleus dililit, dilipat, dan dipadatkan secara tepat menjadi kromosom sehingga dapat masuk ke dalam nukleus. DNA juga diorganisasi sedemikian rupa sehingga segmen-segmen tertentu dapat diakses sesuai kebutuhan oleh jenis sel tertentu.
Tingkat pengorganisasian pertama, atau pengemasan, adalah penggulungan untai DNA di sekitar protein histon. Histon mengemas dan mengatur DNA menjadi unit struktural yang disebut kompleks nukleosom, yang dapat mengontrol akses protein ke wilayah DNA (Gambar 1a). Di bawah mikroskop elektron, penggulungan DNA di sekitar protein histon untuk membentuk nukleosom ini tampak seperti manik-manik kecil pada seutas benang (Gambar 1b). Manik-manik ini (protein histon) dapat bergerak sepanjang benang (DNA) dan mengubah struktur molekul.
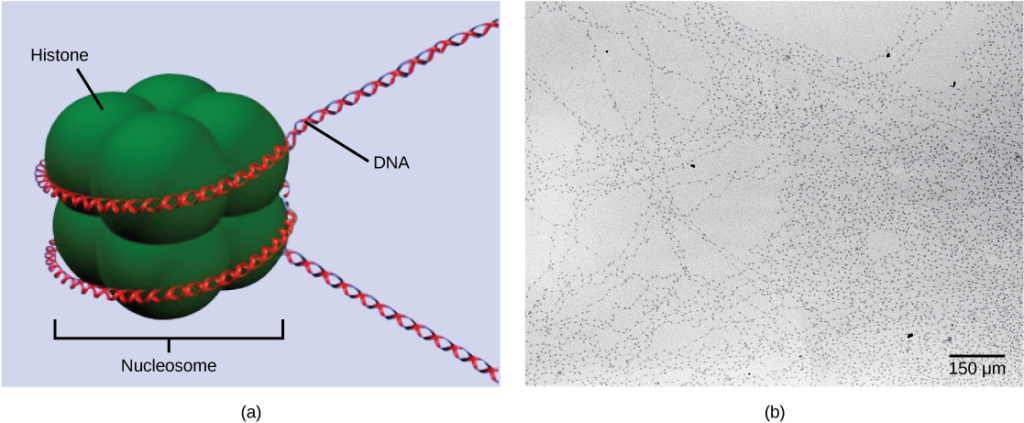
Gambar 1. DNA dilipat di sekitar protein histon untuk menciptakan (a) kompleks nukleosom. Nukleosom ini mengontrol akses protein ke DNA yang mendasarinya. Ketika dilihat melalui mikroskop elektron (b), nukleosom tampak seperti manik-manik pada seutas tali. (kredit “mikrograf”: modifikasi karya Chris Woodcock)
Jika DNA yang mengkode gen tertentu akan ditranskripsikan menjadi RNA, nukleosom yang mengelilingi wilayah DNA tersebut dapat bergeser ke bawah DNA untuk membuka wilayah kromosom spesifik tersebut dan memungkinkan mesin transkripsi (RNA polimerase) untuk memulai transkripsi (Gambar 2). Nukleosom dapat bergerak untuk membuka struktur kromosom guna mengekspos segmen DNA, tetapi melakukannya dengan cara yang sangat terkontrol.
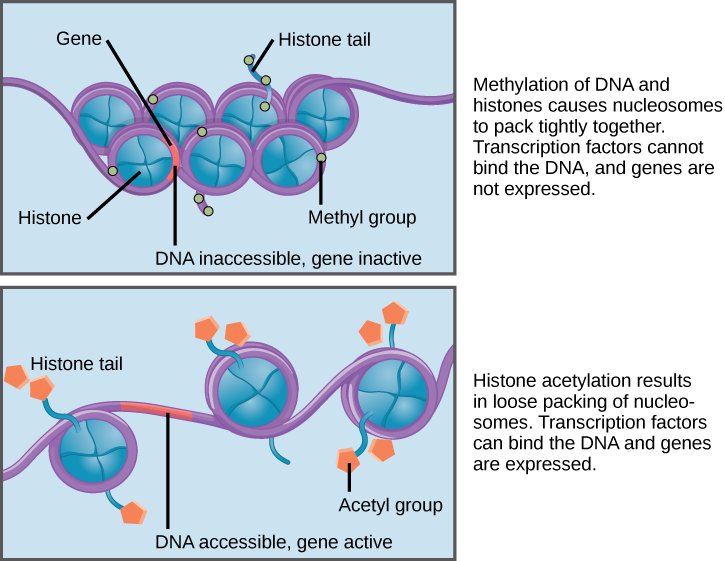
Gambar 2. Nukleosom dapat bergerak di sepanjang DNA. Ketika nukleosom berjarak sangat dekat (atas), faktor transkripsi tidak dapat mengikat dan ekspresi gen dimatikan. Ketika nukleosom berjarak jauh (bawah), DNA terpapar. Faktor transkripsi dapat mengikat, memungkinkan ekspresi gen terjadi. Modifikasi pada histon dan DNA memengaruhi jarak antar nukleosom.
Cara protein histon bergerak bergantung pada sinyal yang ditemukan baik pada protein histon maupun pada DNA. Sinyal-sinyal ini adalah penanda yang ditambahkan ke protein histon dan DNA yang memberi tahu histon apakah suatu wilayah kromosom harus terbuka atau tertutup (Gambar 3 menggambarkan modifikasi pada protein histon dan DNA). Penanda ini tidak permanen, tetapi dapat ditambahkan atau dihilangkan sesuai kebutuhan. Penanda tersebut merupakan modifikasi kimia (gugus fosfat, metil, atau asetil) yang melekat pada asam amino tertentu dalam protein atau pada nukleotida DNA. Penanda tersebut tidak mengubah urutan basa DNA, tetapi mengubah seberapa erat DNA tergulung di sekitar protein histon. DNA adalah molekul bermuatan negatif; oleh karena itu, perubahan muatan histon akan mengubah seberapa erat molekul DNA tergulung. Ketika tidak dimodifikasi, protein histon memiliki muatan positif yang besar; dengan menambahkan modifikasi kimia seperti gugus asetil, muatannya menjadi kurang positif.
Molekul DNA itu sendiri juga dapat dimodifikasi. Hal ini terjadi di dalam wilayah yang sangat spesifik yang disebut pulau CpG. Ini adalah bagian dengan frekuensi tinggi pasangan DNA dinukleotida sitosin dan guanin (CG) yang ditemukan di wilayah promotor gen. Ketika konfigurasi ini ada, anggota sitosin dari pasangan tersebut dapat dimetilasi (gugus metil ditambahkan). Modifikasi ini mengubah cara DNA berinteraksi dengan protein, termasuk protein histon yang mengontrol akses ke wilayah tersebut. Wilayah DNA yang sangat termetilasi (hipermetilasi) dengan histon yang mengalami deasetilasi akan tergulung rapat dan tidak aktif secara transkripsional.
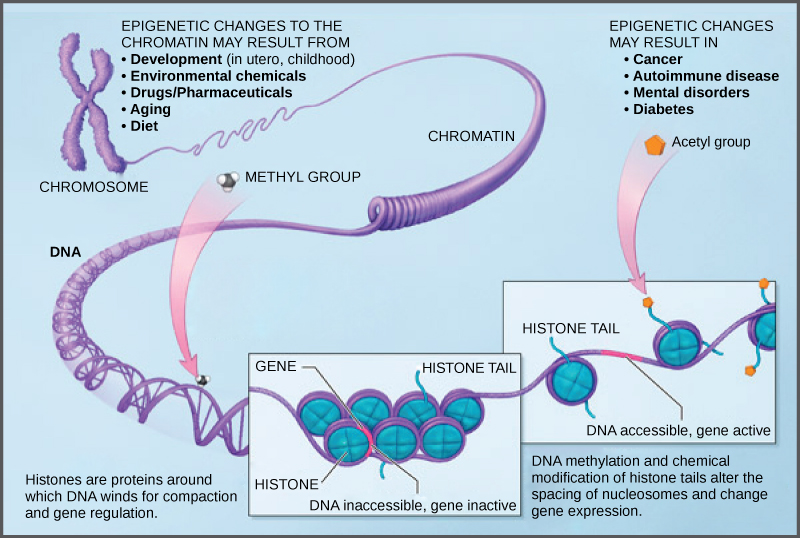
Gambar 3. Protein histon dan nukleotida DNA dapat dimodifikasi secara kimia. Modifikasi tersebut memengaruhi jarak antar nukleosom dan ekspresi gen. (kredit: modifikasi dari karya NIH)
Jenis regulasi gen ini disebut regulasi epigenetik. Epigenetik berarti “di sekitar genetika.” Perubahan yang terjadi pada protein histon dan DNA tidak mengubah urutan nukleotida dan tidak bersifat permanen. Sebaliknya, perubahan ini bersifat sementara (walaupun seringkali bertahan melalui beberapa putaran pembelahan sel) dan mengubah struktur kromosom (terbuka atau tertutup) sesuai kebutuhan. Suatu gen dapat diaktifkan atau dinonaktifkan tergantung pada lokasi dan modifikasi pada protein histon dan DNA. Jika suatu gen akan ditranskripsikan, protein histon dan DNA dimodifikasi di sekitar wilayah kromosom yang mengkode gen tersebut. Ini membuka wilayah kromosom untuk memungkinkan akses bagi RNA polimerase dan protein lain, yang disebut faktor transkripsi, untuk mengikat ke wilayah promotor, yang terletak tepat di hulu gen, dan memulai transkripsi. Jika suatu gen akan tetap dinonaktifkan, atau dibungkam, protein histon dan DNA memiliki modifikasi berbeda yang menandakan konfigurasi kromosom tertutup. Dalam konfigurasi tertutup ini, RNA polimerase dan faktor transkripsi tidak memiliki akses ke DNA dan transkripsi tidak dapat terjadi (Gambar 2).
Seperti sel prokariotik, transkripsi gen pada eukariotik membutuhkan aksi RNA polimerase untuk mengikat sekuens di hulu gen guna memulai transkripsi. Namun, tidak seperti sel prokariotik, RNA polimerase eukariotik membutuhkan protein lain, atau faktor transkripsi, untuk memfasilitasi inisiasi transkripsi. Faktor transkripsi adalah protein yang mengikat sekuens promotor dan sekuens pengatur lainnya untuk mengontrol transkripsi gen target. RNA polimerase sendiri tidak dapat memulai transkripsi pada sel eukariotik. Faktor transkripsi harus terlebih dahulu mengikat daerah promotor dan merekrut RNA polimerase ke lokasi tersebut agar transkripsi dapat dimulai.
Promotor dan Mesin Transkripsi
Gen tersusun sedemikian rupa sehingga memudahkan pengendalian ekspresi gen. Daerah promotor terletak tepat di hulu sekuens pengkodean. Daerah ini bisa pendek (hanya beberapa nukleotida) atau cukup panjang (ratusan nukleotida). Semakin panjang promotor, semakin banyak ruang yang tersedia bagi protein untuk berikatan. Hal ini juga menambah kontrol pada proses transkripsi. Panjang promotor bersifat spesifik gen dan dapat berbeda secara dramatis antar gen. Akibatnya, tingkat kontrol ekspresi gen juga dapat berbeda secara dramatis antar gen. Tujuan promotor adalah untuk mengikat faktor transkripsi yang mengontrol inisiasi transkripsi.
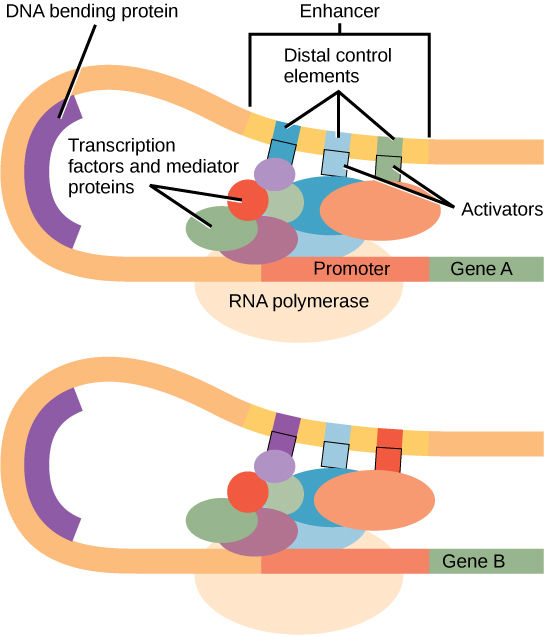
Gambar 1. Enhancer adalah sekuens DNA yang mendorong transkripsi. Setiap enhancer terdiri dari sekuens DNA pendek yang disebut elemen kontrol distal. Aktivator yang terikat pada elemen kontrol distal berinteraksi dengan protein mediator dan faktor transkripsi. Dua gen yang berbeda mungkin memiliki promotor yang sama tetapi elemen kontrol distal yang berbeda, sehingga memungkinkan ekspresi gen yang berbeda.
Di dalam wilayah promotor, tepat di hulu situs awal transkripsi, terdapat kotak TATA. Kotak ini hanyalah pengulangan dinukleotida timin dan adenin (secara harfiah, pengulangan TATA). RNA polimerase berikatan dengan kompleks inisiasi transkripsi, memungkinkan transkripsi terjadi. Untuk memulai transkripsi, faktor transkripsi (TFIID) adalah yang pertama berikatan dengan kotak TATA. Pengikatan TFIID merekrut faktor transkripsi lain, termasuk TFIIB, TFIIE, TFIIF, dan TFIIH ke kotak TATA. Setelah kompleks ini terbentuk, RNA polimerase dapat berikatan dengan sekuens hulunya. Ketika berikatan bersama dengan faktor transkripsi, RNA polimerase difosforilasi. Ini melepaskan sebagian protein dari DNA untuk mengaktifkan kompleks inisiasi transkripsi dan menempatkan RNA polimerase dalam orientasi yang benar untuk memulai transkripsi; protein pembengkok DNA membawa enhancer, yang dapat berada cukup jauh dari gen, bersentuhan dengan faktor transkripsi dan protein mediator (Gambar 1).
Selain faktor transkripsi umum, faktor transkripsi lain dapat berikatan dengan promotor untuk mengatur transkripsi gen. Faktor transkripsi ini berikatan dengan promotor dari sekumpulan gen tertentu. Mereka bukanlah faktor transkripsi umum yang berikatan dengan setiap kompleks promotor, tetapi direkrut ke sekuens spesifik pada promotor gen tertentu. Terdapat ratusan faktor transkripsi dalam sel yang masing-masing berikatan secara spesifik dengan motif sekuens DNA tertentu. Ketika faktor transkripsi berikatan dengan promotor tepat di hulu gen yang dikodekan, hal itu disebut sebagai elemen
cis -acting, karena berada pada kromosom yang sama tepat di sebelah gen. Daerah tempat faktor transkripsi tertentu berikatan disebut situs pengikatan faktor transkripsi. Faktor transkripsi merespons rangsangan lingkungan yang menyebabkan protein menemukan situs pengikatannya dan memulai transkripsi gen yang dibutuhkan.
Peningkat dan Transkripsi
Pada beberapa gen eukariotik, terdapat daerah yang membantu meningkatkan atau memperkuat transkripsi. Daerah-daerah ini, yang disebut enhancer, tidak selalu berdekatan dengan gen yang ditingkatkan. Enhancer dapat terletak di bagian hulu gen, di dalam daerah pengkodean gen, di bagian hilir gen, atau mungkin berjarak ribuan nukleotida.
Daerah penguat (enhancer region) adalah sekuens pengikatan, atau situs, untuk faktor transkripsi. Ketika protein pembengkok DNA mengikat, bentuk DNA berubah (Gambar 1). Perubahan bentuk ini memungkinkan interaksi aktivator yang terikat pada penguat dengan faktor transkripsi yang terikat pada daerah promotor dan RNA polimerase. Meskipun DNA umumnya digambarkan sebagai garis lurus dalam dua dimensi, sebenarnya DNA adalah objek tiga dimensi. Oleh karena itu, sekuens nukleotida yang berjarak ribuan nukleotida dapat melipat dan berinteraksi dengan promotor tertentu.
Mematikan Gen: Represor Transkripsi
Seperti sel prokariotik, sel eukariotik juga memiliki mekanisme untuk mencegah transkripsi. Represor transkripsi dapat mengikat daerah promotor atau enhancer dan memblokir transkripsi. Seperti aktivator transkripsi, represor merespons rangsangan eksternal untuk mencegah pengikatan faktor transkripsi pengaktif.
Kontrol Pasca-Translasi Ekspresi Gen
RNA ditranskripsikan, tetapi harus diproses menjadi bentuk matang sebelum translasi dapat dimulai. Pemrosesan setelah molekul RNA ditranskripsikan, tetapi sebelum diterjemahkan menjadi protein, disebut modifikasi pasca-transkripsi. Seperti halnya tahap epigenetik dan transkripsi dalam pemrosesan, langkah pasca-transkripsi ini juga dapat diatur untuk mengontrol ekspresi gen dalam sel. Jika RNA tidak diproses, dipindahkan, atau diterjemahkan, maka tidak akan ada protein yang disintesis.
Penyambungan RNA, tahap pertama dari kontrol pasca-transkripsi.
Pada sel eukariotik, transkrip RNA sering mengandung daerah yang disebut intron, yang dihilangkan sebelum translasi. Daerah RNA yang mengkode protein disebut ekson (Gambar 1). Setelah molekul RNA ditranskripsikan, tetapi sebelum meninggalkan nukleus untuk diterjemahkan, RNA diproses dan intron dihilangkan melalui penyambungan (splicing).
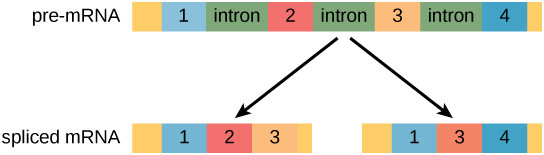
Gambar 1. Pra-mRNA dapat mengalami penyambungan alternatif untuk menghasilkan protein yang berbeda.
Penyambungan RNA Alternatif
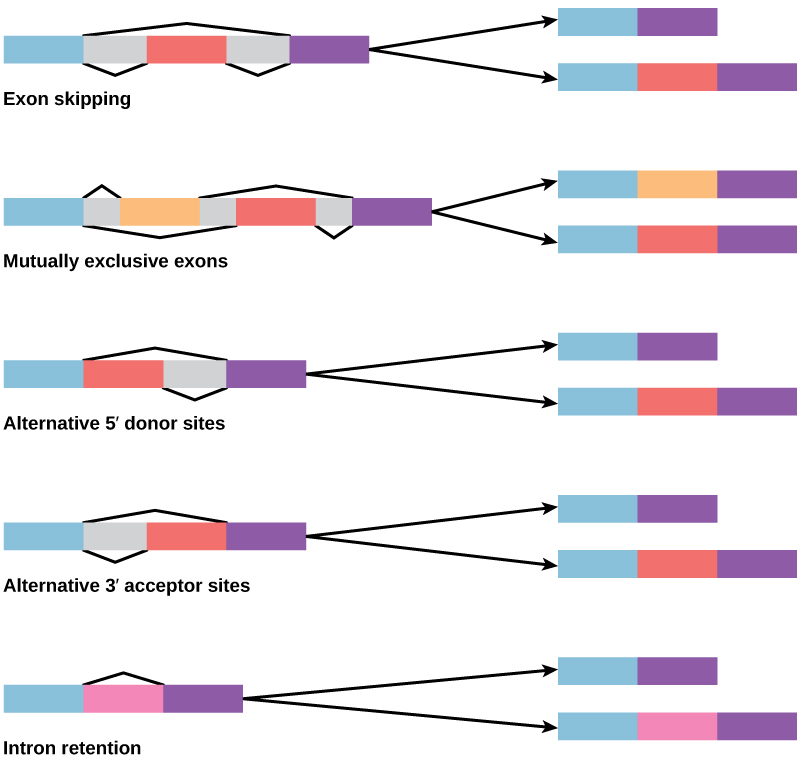
Gambar 2. Ada lima mode dasar penyambungan alternatif.
Pada tahun 1970-an, gen pertama kali diamati yang menunjukkan penyambungan RNA alternatif. Penyambungan RNA alternatif adalah mekanisme yang memungkinkan produk protein yang berbeda dihasilkan dari satu gen ketika kombinasi intron yang berbeda, dan terkadang ekson, dihilangkan dari transkrip (Gambar 2). Penyambungan alternatif ini dapat terjadi secara acak, tetapi lebih sering dikendalikan dan bertindak sebagai mekanisme regulasi gen, dengan frekuensi alternatif penyambungan yang berbeda dikendalikan oleh sel sebagai cara untuk mengontrol produksi produk protein yang berbeda di sel yang berbeda atau pada tahap perkembangan yang berbeda. Penyambungan alternatif sekarang dipahami sebagai mekanisme regulasi gen yang umum pada eukariota; menurut satu perkiraan, 70 persen gen pada manusia diekspresikan sebagai beberapa protein melalui penyambungan alternatif.
Bagaimana penyambungan alternatif dapat berevolusi? Intron memiliki sekuens pengenalan awal dan akhir; mudah untuk membayangkan kegagalan mekanisme penyambungan untuk mengidentifikasi ujung intron dan malah menemukan ujung intron berikutnya, sehingga menghilangkan dua intron dan ekson di antaranya. Faktanya, ada mekanisme yang mencegah penghilangan intron tersebut, tetapi mutasi kemungkinan besar akan menyebabkan kegagalannya. “Kesalahan” semacam itu kemungkinan besar akan menghasilkan protein yang tidak berfungsi. Memang, penyebab banyak penyakit genetik adalah penyambungan alternatif, bukan mutasi dalam sekuens. Namun, penyambungan alternatif akan menciptakan varian protein tanpa kehilangan protein asli, membuka kemungkinan adaptasi varian baru tersebut terhadap fungsi baru. Duplikasi gen telah memainkan peran penting dalam evolusi fungsi baru dengan cara yang serupa, yaitu dengan menyediakan gen yang dapat berevolusi tanpa menghilangkan protein asli yang fungsional.
Pengendalian Stabilitas RNA
Sebelum mRNA meninggalkan nukleus, ia diberi dua “tutup” pelindung yang mencegah ujung untai terdegradasi selama perjalanannya. Tutup 5′ , yang ditempatkan di ujung 5′ mRNA, biasanya terdiri dari molekul guanosin trifosfat (GTP) termetilasi. Ekor poli-A , yang melekat pada ujung 3′, biasanya terdiri dari serangkaian nukleotida adenin. Setelah RNA diangkut ke sitoplasma, lamanya waktu RNA berada di sana dapat dikontrol. Setiap molekul RNA memiliki masa hidup yang ditentukan dan meluruh pada laju tertentu. Laju peluruhan ini dapat memengaruhi jumlah protein dalam sel. Jika laju peluruhan meningkat, RNA tidak akan berada di sitoplasma selama itu, sehingga memperpendek waktu terjadinya translasi. Sebaliknya, jika laju peluruhan menurun, molekul RNA akan berada di sitoplasma lebih lama dan lebih banyak protein dapat ditranslasikan. Laju peluruhan ini disebut sebagai stabilitas RNA. Jika RNA tersebut stabil, maka RNA tersebut akan terdeteksi dalam jangka waktu yang lebih lama di dalam sitoplasma.
Pengikatan protein ke RNA dapat memengaruhi stabilitasnya. Protein, yang disebut protein pengikat RNA , atau RBP, dapat mengikat ke wilayah RNA tepat di hulu atau hilir wilayah pengkodean protein. Wilayah-wilayah dalam RNA yang tidak diterjemahkan menjadi protein ini disebut wilayah yang tidak diterjemahkan , atau UTR. Wilayah ini bukanlah intron (intron telah dihilangkan di dalam nukleus). Sebaliknya, ini adalah wilayah yang mengatur lokalisasi mRNA, stabilitas, dan translasi protein. Wilayah tepat sebelum wilayah pengkodean protein disebut 5′ UTR , sedangkan wilayah setelah wilayah pengkodean disebut 3′ UTR (Gambar 3). Pengikatan RBP ke wilayah-wilayah ini dapat meningkatkan atau menurunkan stabilitas molekul RNA, tergantung pada RBP spesifik yang mengikat.
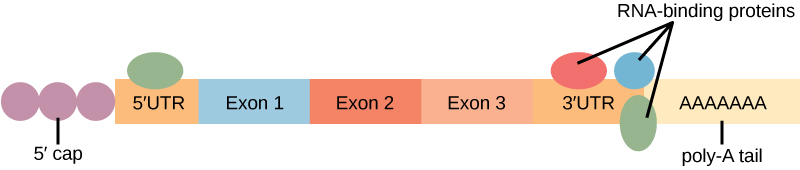
Gambar 3. Daerah pengkodean protein mRNA diapit oleh daerah tak diterjemahkan 5′ dan 3′ (UTR). Keberadaan protein pengikat RNA pada UTR 5′ atau 3′ memengaruhi stabilitas molekul RNA.
Stabilitas RNA dan mikroRNA
Selain RBP yang mengikat dan mengontrol (meningkatkan atau menurunkan) stabilitas RNA, elemen lain yang disebut mikroRNA dapat mengikat molekul RNA. MikroRNA , atau miRNA, adalah molekul RNA pendek yang hanya berukuran 21–24 nukleotida. miRNA dibuat di nukleus sebagai pre-miRNA yang lebih panjang. Pre-miRNA ini dipotong menjadi miRNA matang oleh protein yang disebut dicer . Seperti faktor transkripsi dan RBP, miRNA matang mengenali sekuens spesifik dan mengikat RNA; namun, miRNA juga berasosiasi dengan kompleks ribonukleoprotein yang disebut kompleks penekan RNA (RISC) . RISC mengikat bersama dengan miRNA untuk mendegradasi mRNA target. Bersama-sama, miRNA dan kompleks RISC dengan cepat menghancurkan molekul RNA.
Regulasi Gen Eukariot
1) Tinjauan umum
Manusia merupakan organisme multiseluler yang tersusun atas berbagai macam sel, diantaranya adalah sel darah merah, sel saraf, dan sel otot yang masing-masing memiliki fungsi dan bentuk yang berbeda. Namun demikian, meskipun setiap sel tersebut memiliki fungsi dan bentuk yang berbeda, mereka tetap memiliki susunan gen yang sama karena mulanya sel-sel tersebut berasal dari satu sel yang sama yaitu zigot yang kemudian berdiferensiasi membentuk berbagai macam sel. Perbedaan fungsi dan bentuk dari setiap sel tersebut disebabkan oleh peristiwa regulasi ekspresi gen dimana terdapat gen yang diaktifkan dan terdapat gen yang dinonaktifkan.
2) Karakteristik Ekspresi gen pada eukariotik
Ekspresi gen pada eukariotik bersifat lebih rumit dibandingkan dengan prokariotik. Hal ini terjadi karena sel eukariotik memiliki sistem sel yang kompartemen (organelnya dipisahkan oleh membran). Pembagian sel eukariotik melalui sistem kompartemen ini menyebabkan terjadinya pemisahan tingkatan peristiwa ekspresi gen, seperti yang ditunjukkan pada (Gambar 1) Proses transkripsi DNA menjadi RNA terjadi di dalam nukleus dimana mRNA yang dihasilkan akan diekspor ke sitoplasma untuk berikatan dengan ribosom. Setelah berikatan dengan ribosom, mRNA ini ditranslasikan menjadi polipeptida. Pemisahan tingkatan peristiwa ekspresi gen ini memungkinkan regulasi ekspresi gen dapat terjadi di tempat yang berbeda dimana regulasi dapat terjadi di dalam nukleus pada tingkat DNA atau RNA, atau di dalam sitoplasma pada tingkat RNA atau polipeptida.
Gambar 1. Peristiwa regulasi ekspresi gen pada eukariotik yang dapat terjadi pada berbagai tahapan (Snustad & Simmons, 2020)
2) Kontrol Sitolasmik Stabilitas mRNA
a. Suhu: Gen Heat-Shock
Memiliki kesamaan dengan prokariotik, regulasi ekspresi gen pada eukariotik juga dapat dipengaruhi oleh lingkungan. Salah satunya adalah gen HSP70 pada D. melanogaster yang dapat diekspresikan ketika temperatur lingkungan yang tinggi. Ketika temperatur lingkungan yang normal, gen HSP70 normalnya tidak diekspresikan. Kinerja dari gen HSP70 sendiri diatur oleh HSE (Heat-shock response elements) dimana secara khusus, gen HSP70 akan ditranskripsikan apabila daerah HSE berikatan dengan HTSF (Heat-shock transcription factor). Ketika temperatur lingkungan berada pada kondisi normal, HSTF akan bersifat inaktif. Namun demikian apabila temperatur lingkungan tinggi, HSTF akan bersifat aktif (Gambar 2)
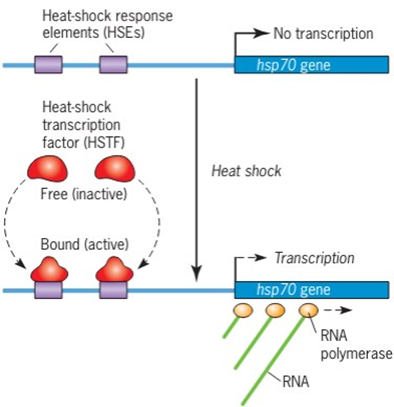
Gambar 2. Induksi transkripsi gen HSP70 D. melanogaster oleh kejut panas
(Snustad & Simmons, 2020)
3) Molekul Sinyal: Gen yang Merespon Hormon
- Hormon steroid
Hormon steroid terdiri dari steroid yang merupakan turunan dari lipid. Sel eukariotik tersusun oleh membran sel yang juga tersusun atas fosfolipid sehingga steroid dapat mudah terlarut dalam lipid dan mampu melewati membran sel (Gambar 3). Setelah memasuki sitoplasma, hormon steroid akan menempel pada protein reseptor. Akibatnya, hormon steroid dan protein reseptor akan membentuk kompleks yang memasuki inti sel melalui membran ini. Kompleks tersebut nantinya akan menempel pada DNA dan apabila kompleks hormon steroid serta protein reseptor sudah menempel pada DNA, hal ini akan menginduksi DNA untuk ditranskripsikan. Hasil transkripsi nantinya akan dibawa ke sitoplasma untuk di translasikan.
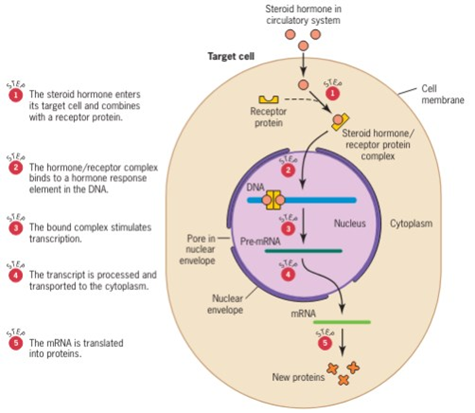
Gambar 3. Regulasi ekspresi gen pada eukariotik yang dipicu oleh hormon steroid
(Snustad & Simmons, 2020)
- Hormon peptida
Hormon peptida memiliki ukuran yang lebih besar dan tidak terlarut dalam lemak sehingga ia tidak dapat masuk ke dalam sitoplasma. Apabila hormon peptida ingin memasuki membran sel, maka ia harus berikatan dengan protein reseptor pada membran sel terlebih dahulu. Ketika sudah berhasil berikatan, protein di bawah protein reseptor akan mengalami aktivasi, sehingga ia akan menginduksi molekul persinyalan untuk masuk ke dalam inti sel (Gambar 4). Setelah molekul persinyalan berhasil memasuki inti sel, ia akan menginduksi faktor transkripsi untuk menempel pada DNA. Dengan demikian, promoter akan menempel dengan polimerase dan DNA dapat ditranskripsikan menjadi RNA untuk kemudian di transpor ke dalam ribosom untuk di translasikan.
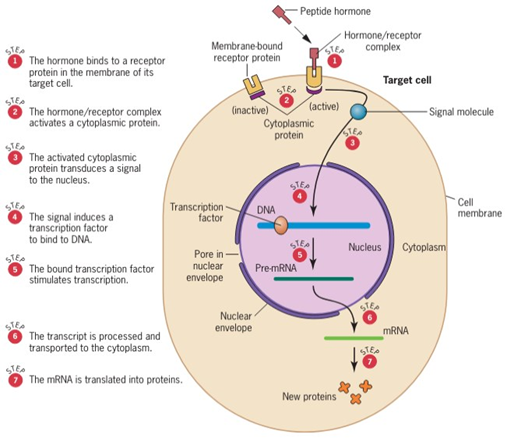
Gambar 4. Regulasi ekspresi gen pada eukariotik yang dipicu oleh hormon peptida
(Sumber: Snustad & Simmons, 2020)
Kontrol molekuler serta kontrol posttranscriptional regulation oleh RNA interference Enhancer merupakan segmen pada DNA yang berfungsi untuk meregulasi ekspresi gen lainnya. Enhancer memiliki fungsi meningkatkan laju ekspresi gen lainnya. Contohnya adalah gen kuning (yellow gene) pada Drosophila yang bertanggung jawab untuk pigmentasi di beberapa bagian tubuh (sayap, kaki, punggung, dan perut (Gambar 5).
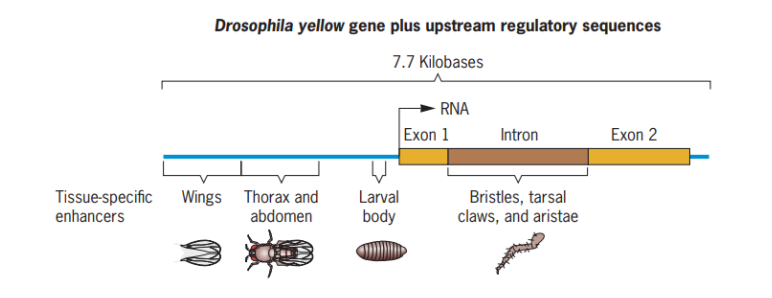
Gambar 5. Enhancer spesifik pada jaringan dari gen kuning Drosophila
(Snustad & Simmons, 2020)
Enhancer merupakan salah satu komponen di dalam sel yang berperan meregulasi ekspresi gen lainnya. Fungsi enhancer adalah meningkatkan laju ekspresi gen lain. Contohnyaadalah enhancer yang mengatur ekspresi gen yellow pada Drosophila. Adapun skema gambar dari gen yellow dapat dilihat pada Gambar 5.
Pada gen yellow Drosophila, terdapat ekson 1, intron, dan ekson 2 dengan keberadaan intron di tengah-tengah ekson 1 dan 2. Pada gen yellow Drosophila, enhancer ada pada sayap, thorax dan abdomen, serta tubuh larva yang memiliki fungsi sama yaitu meningkatkan laju transkripsi. Namun demikian, tidak semua enhancer ini bekerja bersama-sama untuk meningkatkan laju transkripsi. Enhancer dikatakan sebagai spesifik jaringan, yang artinya pada jaringan tertentu, enhancer (contoh: A) yang bekerja, sedangkan jaringan pada lain enhancer (contoh: B) yang bekerja. Sama seperti enhancer pada gen yellow ini, apabila enhancer yang berada di sel-sel penyusun sayap, enhancer yang bekerja meningkatkan laju transkripsi dari gen yellow adalah enhancer di daerah sayap. Apabila pada sel-sel penyusun thorax dan abdomen, maka enhancer yang bekerja adalah enhancer thorax dan abdomen.
Apabila enhancer berperan meningkatkan laju transkripsi gen yellow ketika Drosophila masih berbentuk larva maka enhancer yang bekerja adalah di daerah enhancer tubuh larva. Apabila gen yellow yang ditingkatkan laju transkripsinya merupakan gen-gen yang ditemukan di sel-sel penyusun bristle, tarsal, claws, dan aristae dari bagian tubuh Drosophila, maka enhancer yang bekerja adalah enhancer yang ditemukan di intron.
Enhancer memiliki sifat yang menarik bahwa dia dikatakan independensi lokasi artinya enhancer itu dapat ditemukan di beberapa lokasi di dalam suatu gen. Enhancer juga dikatakan sebagai independensi orientasi (arah). Sebagai contoh enhancer dengan urutan A-U-G-C-C-C. yang ketika dipotong dan oritensasi nya dibalik, urutannya menjadi C-C-C-G-U-A. Namun demikian, meski urutan dari enhancer tersebut terbalik, dia masih dapat bekerja menjadi enhancer dan mampu meningkatkan laju transkripsi dari gen-gen yang diatur. Selain itu, enhancer masih dapat meregulasi gen meskipun jaraknya sangat jauh sampai ribuan basa jauhnya dari gen yang diatur.
1) Cara enhancer dapat meregulasi ekspresi gen yang jaraknya jauh
Enhancer akan bekerja ketika terdapat protein aktivator yang menempel dengannya. Kemudian, DNA akan melengkung dan akan mendekati promotor sehingga kompleks enhancer-protein tersebut akan mendekati promotor. Ketika kompleks enhancer-protein dan promotor sudah berikatan, kompleks ini akan mempermudah faktor transkripsi hingga RNA polimerase untuk berikatan dengan promotor. Dengan demikian, dapat disimpulkan bahwa enhancer dapat mempermudah penempelan RNA polimerase di promotor dari gen yang dia atur. begitu Selain itu, enhancer juga dapat memfasilitasi atau mempermudah penempelan faktor-faktor transkripsi dengan promotor dari gen yang diatur. Oleh karena faktor transkripsi terfasilitasi untuk menempel dengan promotor dari gen yang diatur, maka RNA polimerase pada akhirnya juga dapat menempel pada promotor di gen tersebut. Adapun mekanisme cara kerja enhancer untuk dapat meregulasi ekspresi gen yang jaraknya jauh dapat dilihat pada Gambar 6.
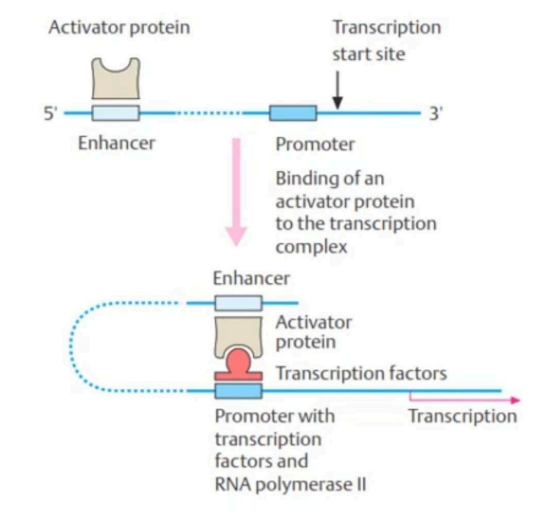
Gambar 6. Mekanisme cara kerja enhancer untuk meregulasi gen yang jaraknya jauh
(Passarge, 2007)
Selain enhancer, faktor transkripsi juga berperan dalam mekanisme regulasi ekspresi gen. Faktor transkripsi terdiri dari 2 macam yaitu, faktor transkripsi dasar dan faktor transkripsi khusus. Faktor-faktor transkripsi yang dapat menarik kehadiran RNA polimerase untuk mengikat dengan promotor disebut dengan faktor transkripsi dasar. Sedangkan, faktor-faktor transkripsi yang dapat berikatan dengan enhancer seperti pada Gambar 6. merupakan faktor transkripsi khusus.
Faktor transkripsi sebetulnya merupakan protein dimana pada dasarnya protein tersusun atas beberapa motif atau struktur khusus dari rantai polipeptida yang menyusun protein. Adapun beberapa susunan motif dari faktor transkripsi dapat dilihat pada Gambar 7. Komposisi dari keberadaan motif-motif tersebut di dalam sel dapat mengatur regulasi ekspresi gen tertentu. Contohnya, gen A hanya dapat ditranskripsikan apabila ia menempel dengan faktor transkripsi yang tersusun atas zinc-finger motif. Apabila suatu sel memiliki komposisi zinc-finger motif yang rendah, maka laju transkripsi dari gen A juga akan rendah. Namun demikian, apabila gen A dari sel lain memiliki laju transkripsi yang tinggi, maka kandungan zinc-finger motif juga akan tinggi. Dengan demikian, apa jumlah motif semakin tinggi, maka akan semakin mudah pula terbentuknya faktor transkripsi, sehingga faktor transkripsi akan mudah menempel dengan promotor. Dalam konteks tersebut, gen A akan memiliki laju transkripsi yang juga tinggi.
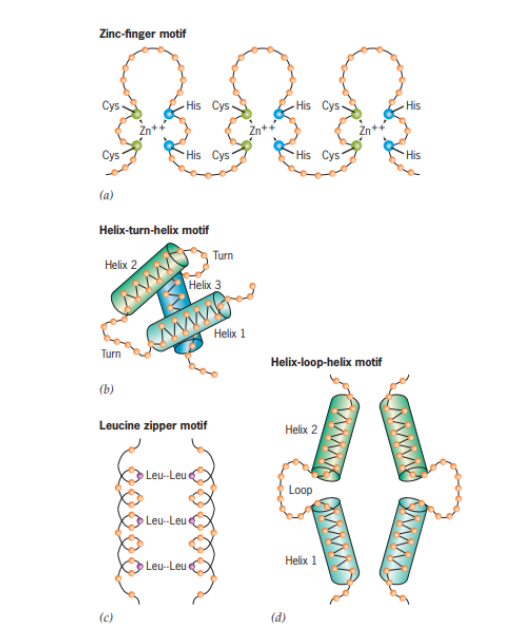
Gambar 7. Susunan motif dari faktor transkripsi
(Sumber: Snustad & Simmons, 2020)
Mekanisme regulasi ekspresi gen pada eukariotik tidak hanya diatur pada level transkripsi. Artinya, meskipun DNA sudah ditranskripsikan, hasil transkripsi ini juga dapat diatur regulasinya yaitu dapat ditekan atau dilanjutkan hingga level translasi. Salah satu komponen yang terlibat dalam mekanisme regulasi ekspresi gen eukariotik pada mekanisme pasca transkripsi adalah RNAi (RNA interference). RNAi merupakan RNA yang dapat menempel dengan mRNA tertentu sehingga RNA tersebut tidak dapat dilanjutkan hingga level translasi.
RNAi berasal dari molekul RNA unting ganda yang berukuran besar atau panjang, dimana lebih tepatnya kebanyakan dihasilkan oleh gen mir. Gen mir akan ditranskripsikan menjadi RNA unting tunggal. Namun demikian, terdapat beberapa daerah yang basanya komplementer sehingga unting RNA tunggal yang awalnya komplementer tersebut dapat membentuk loop atau lekukan hingga menjadi RNA unting ganda seperti pada Gambar 8a. Kenudian, terdapat enzim lain yang akan memotong loop tersebut sehingga akan membentuk RNA unting ganda tanpa loop seperti yang ditunjukkan pada Gambar 8b.
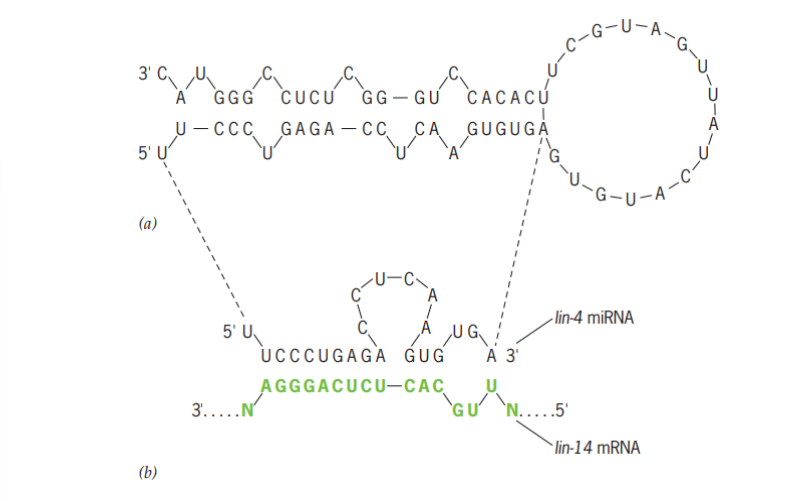
Gambar 8. Regulasi ekspresi gen oleh RNAi
(Sumber: Snustad & Simmons. 2021)
Adapun mekanisme jalur RNAi dapat dilihat pada Gambar 9.
RNA unting ganda tersebut kemudian dipotong oleh dicer (1) menjadi bagian-bagian yang lebih kecil. Setelah molekul RNA unting ganda tersebut dipotong menjadi beberapa bagian yang kecil, bagian tersebut nantinya dapat ditempeli oleh protein khusus, yang dikenal sebagai ribonucleoprotein (2). Setelah itu, RNA yang mulanya untai ganda akan dijadikan sebagai unting tunggal (3). Kompleks tersebut disebut dengan RISC yaitu RNA-induced Silencing Complex yang tersusun atas protein dan unting RNA tunggal. Setelah RISC terbentuk, RNA tersebut sudah dapat bekerja sebagai RNAi dimana ia akan menempel pada mRNA target (4 & 5). Ketika sudah menempel pada mRNA target, terdapat 2 kemungkinan, yaitu yang pertama adalah penempelan sempurna (6A). Hal ini terjadi ketika pertama hingga terakhir dari RNAi dapat menempel dengan sempurna dengan basa-basa yang ada pada mRNA targetnya. Kemungkinan yang kedua adalah hanya beberapa absa yang sempurna tempelannya sedangkan sebagiannya tidak komplemen pada mRNA target sehingga membentuk semacam lengkungan (6B). Ketika RNAi menempel sempurna, RNAi akan bertindak sebagai siRNA (small interfering RNA). Dengan siRNA menempel dengan mRNA. Kemudian, mRNA akan di degradasi atau di potong-potong. Namun demikian, apabila RNAi membentuk pasangan yang tidak sempurna, akan terbentuk miRNA (micro RNA). Ketika miRNA menempel dengan mRNA, RNA tidak akan ditranslasi karena kehadiran miRNA akan mengganggu ribosom untuk membantu melakukan translasi. Dengan demikian, baik dalam siRNA atau miRNA, keberadaan RNAi akan menghambat atau menekan peristiwa translasi dari mRNA.
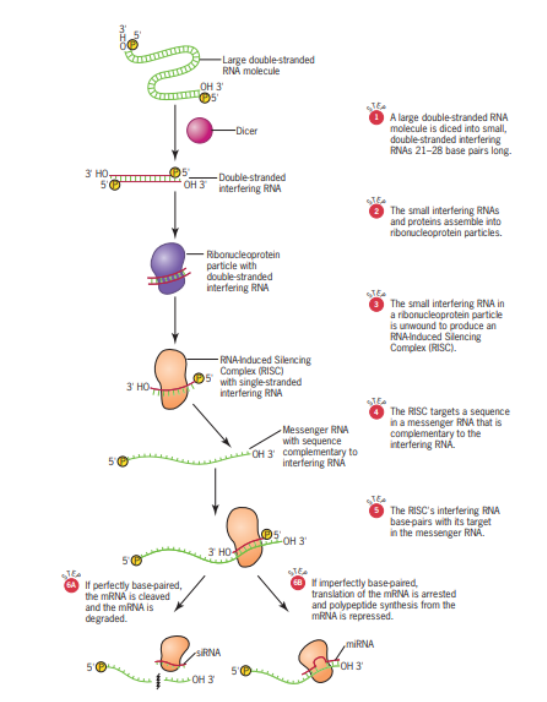
Gambar 9. Jalur RNAi
(Sumber: Snustad & Simmons, 2021)
Asetilasi Histon, Metilasi DNA, dan Imprinting
Gambar 10 merupakan gambar dari kromosom yang bisa ditemukan di dalam inti sel eukariotik Dalam struktur kromosom, ternyata terdapat daerah-daerah yang sangat memampat dan terdapat daerah-daerah yang lebih renggang. Daerah-daerah yang lebih mampat atau lebih rapat dikenal sebagai heterokromatin, sedangkan daerah-daerah yang lebih renggang dikenal sebagai eukromatin. Gen-gen yang secara aktif ditranskripsikan atau diekspresikan merupakan gen-gen yang berada di daerah eukromatin atau daerah yang lebih renggang, sedangkan di daerah heterokromatin, segmen-segmen DNA atau gen-gen di daerah tersebut diinaktifkan atau laju transkripsinya lebih rendah. Peristiwa ini merupakan salah satu contoh bagaimana gen-gen di suatu sel dapat diinaktifkan sehingga ketika gen-gen tersebut berada di daerah heterokromatin, maka gen-gen tersebut menjadi inaktif. Lebih lanjut, mungkin saja awalnya suatu daerah itu merupakan eukromatin yang kemudian terjadi modifikasi sehingga eukromatin itu berubah menjadi heterokromatin atau yang awalnya renggang kemudian menjadi memampat sehingga awalnya gen-gen tersebut dapat diaktifkan atau dapat diekspresikan, namun setelah memampat
menjadi heterokromatin, maka gen-gen tadi menjadi inaktif.
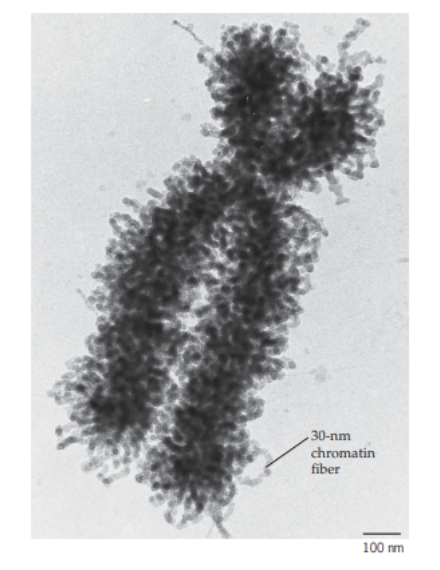
Gambar 10. Tampilan kromosom pada manusia
(Sumber: Snustad & Simmons, 2021)
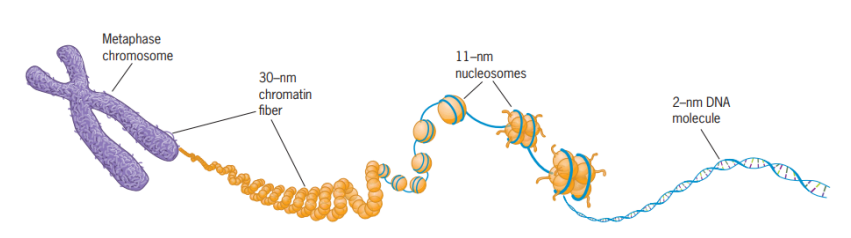
Kromosom sebetulnya tersusun atas benang kromatin yang memadat (dapat dibagi menjadi daerah eukromatin dan daerah heterokromatin). Kromatin ini sebenarnya tersusun atas dua komponen utama, yaitu DNA dan protein. DNA meliliti protein-protein yang kita kenal sebagai protein histon yang ditunjukkan dengan komponen kuning pada Gambar 31, sedangkan DNA-nya ditunjukkan dengan unting berwarnanya biru pada Gambar 11.
Gambar 31. Komponen kromosom
(Sumber: Snustad & Simmons, 2021)
Apabila terjadi modifikasi baik pada DNA ataupun protein histonnya, akan juga mempengaruhi laju transkripsi atau ekspresi genetik dari gen tersebut. Contohnya, pada peristiwa chromatin remodeling. Chromatin remodeling merupakan perubahan struktur kromosom yang dapat dikatakan menjadi lebih terbuka strukturnya sehingga komponen-komponen yang terlibat dalam transkripsi dapat masuk ke daerah tersebut. Sebagai contoh, ketika DNA meliliti histon dan memampat, maka komponen-komponen yang terlibat dalam transkripsi itu sulit untuk masuk. Dengan chromatin remodeling memungkinkan peralatan transkripsi hadir dan masuk di daerah segmen DNA tersebut sehingga segmen DNA tersebut dapat ditranskripsikan.
Asetilasi Histon
Salah satu peristiwa dalam chromatin remodeling adalah asetilasi histon atau penempelan asetil pada protein histon (Gambar 12). Oleh karena protein tersusun atas rantai asam amino, maka pada Gambar 12, terdapat rantai yang tersusun atas banyak asam amino. Asetilasi histon mampu meningkatkan laju transkripsi dari DNA tersebut sehingga misalkan aterda segmen DNA segmen DNA itu meliliti histon dan ketika histon tersebut mengalami asetilasi atau penempelan gugus asetil pada daerah histon tersebut (khususnya di asam amino lisin), maka laju transkripsi segmen DNA tersebut akan meningkat.

Gambar 12. Asetilasi histon
(Snustad & Simmons, 2021)
Sebaliknya ketika DNA mengalami metilasi, laju transkripsi dari segmen DNA tersebut akan ditekan. Metilasi merupakan penempelan gugus metil (CH3) pada basa sitosin (Gambar 13). Dengan demikian, ketika histon terasetilasi, laju transkripsi akan meningkat sedangkan ketika sitosin termetilasi laju transkripsinya menurun.
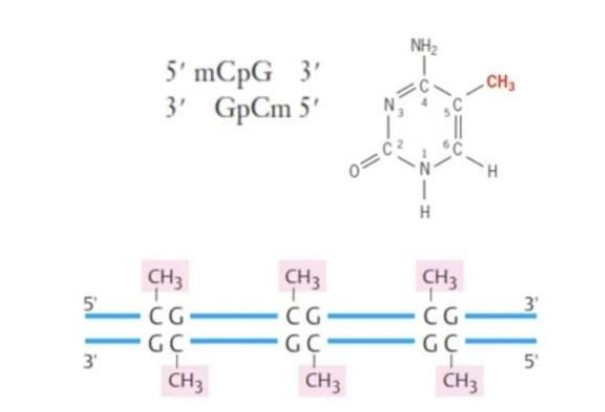
Gambar 13. Metilasi DNA
(Passarge, 2007)
Imprinting
Imprinting merupakan peristiwa yang mengakibatkan suatu pasangan gen hanya mentranskripsikan salah satu alelnya saja dan transkripsi salah satu alel tersebut berkaitan dengan dari mana alel tersebut berasal. Dalam konteks ini, alel tersebut dapat berasal dari orang tua, yaitu laki-laki-perempuan atau betina-jantan (Gambar 14).
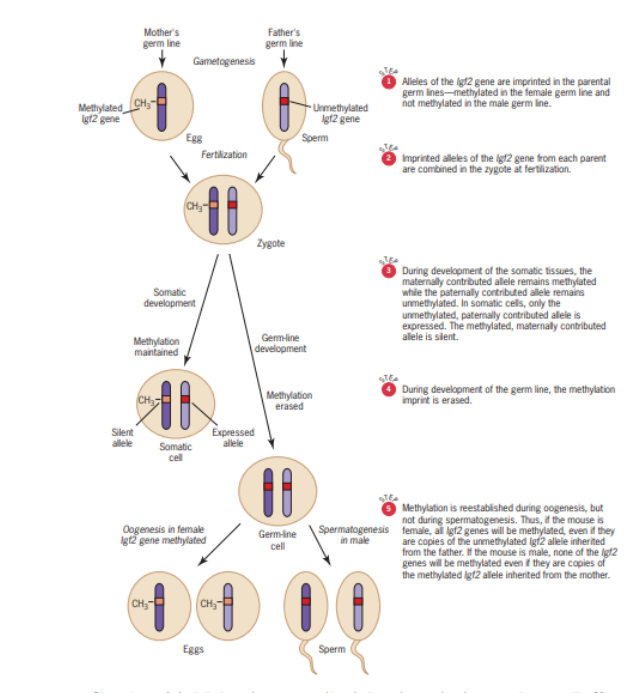
Gambar 14. Mekanisme metilasi dan imprinting pada gen Igf2
(Sumber: Snustad & Simmons, 2021)
Contohnya, gen Igf2 dalam tubuh memiliki jumlah sepasang sehingga terdapat dua alel yaitu satu dari ibu sedangkan yang satu lainnya berasal dari ayah. Dalam konteks ini, ibu menghasilkan sel telur, sedangkan ayah menghasilkan sel sperma yang keduanya baik sel telur maupun sperma membawa alel Igf2.
Ternyata Igf2 dari ibu atau sel telur mengalami metilasi dan ketika suatu gen mengalami metilasi, maka gen itu tidak diaktifkan. Dalam Gambar 14, gen Igf2 pada sel telur tidak diaktifkan, berbeda dengan sel sperma yang tidak mengalami metilasi pada Igf2-nya. Ketika terjadi fertilisasi, kromosom ibu dan ayah berpasangan sehingga Igf2-nya pun menjadi memiliki dua alel, yaitu satu dari ibu dan satu lainnya dari ayah.
Namun demikian, Igf2 dari ibu termetilasi, sedangkan Igf2 pada ayah tidak termetilasi. Dengan demikian, gen yang diekspresikan hanya alel Igf2 yang berasal dari ayah. Ketika zigot ini membelah dan berkembang menjadi sel somatik, metilasi tersebut akan dipertahankan. Lebih lanjut, alel dari ibu selalu termetilasi, sehingga tidak diaktifkan. Sedangkan alel dari ayah itu tidak termetilasi sehingga selalu diekspresikan secara normal.
Namun demikian, ketika zigot ini berkembang menjadi calon sperma dan sel telur, maka metilasi tersebut dibuang. Ketika zigot tersebut merupakan laki-laki yang akan menghasilkan sperma, spermanya tetap tidak akan termetilasi. Namun demikian, apabila zigot ini merupakan perempuan yang akan menghasilkan sel telur, sel telur yang dihasilkan akan langsung termetilasi kembali.
Sumber Rujukan:
- Snustad & Simmons. (2020). Principles of genetics (7th ed.). Hoboken, NJ: John Wiley & Sons.
- Snustad & Simmons. (2021). Principles of genetics (8th ed.). Hoboken, NJ: John Wiley & Sons.
- Passarge, E. (2007). Color atlas of genetics (3rd ed.). Stuttgart: Thieme Verlag.
- Gardner, E. J. (1991). Principles of genetics (8th ed.). New York, NY: John Wiley & Sons.
Ringkasan materi:
- Regulasi ekspresi gen adalah proses pengendalian aktivitas gen dalam menghasilkan RNA dan protein.
- Menentukan kapan, di mana, dan seberapa banyak suatu gen diekspresikan.
- Memungkinkan sel yang memiliki DNA sama menghasilkan fungsi yang berbeda (diferensiasi sel).
- Konsep dasar regulasi gen dijelaskan melalui model operon oleh François Jacob dan Jacques Monod.
- Pada prokariotik, regulasi terutama terjadi pada tingkat transkripsi melalui sistem operon (misalnya operon lac dan operon trp).
- Pada eukariotik, regulasi lebih kompleks dan terjadi pada berbagai tingkat.
- Tingkat regulasi meliputi:
- Regulasi transkripsi (promoter, enhancer, faktor transkripsi).
- Regulasi pasca-transkripsi (splicing, capping, polyadenilasi, stabilitas mRNA).
- Regulasi translasi (kontrol sintesis protein).
- Regulasi pasca-translasi (modifikasi protein seperti fosforilasi dan ubiquitinasi).
- Regulasi epigenetik (metilasi DNA dan modifikasi histon).
- Regulasi ekspresi gen berperan dalam perkembangan embrio, adaptasi terhadap lingkungan, dan respons seluler.
- Gangguan regulasi ekspresi gen dapat menyebabkan penyakit seperti kanker dan kelainan metabolik.
Ringkasan: Regulasi Gen Epigenetik Eukariotik
Pada sel eukariotik, tahap pertama pengendalian ekspresi gen terjadi pada tingkat epigenetik. Mekanisme epigenetik mengontrol akses ke wilayah kromosom untuk memungkinkan gen diaktifkan atau dinonaktifkan. Mekanisme ini mengontrol bagaimana DNA dikemas ke dalam nukleus dengan mengatur seberapa erat DNA dililitkan di sekitar protein histon. Penambahan atau penghapusan modifikasi kimia (atau penanda) pada protein histon atau DNA memberi sinyal kepada sel untuk membuka atau menutup wilayah kromosom. Oleh karena itu, sel eukariotik dapat mengontrol apakah suatu gen diekspresikan dengan mengontrol aksesibilitas ke faktor transkripsi dan pengikatan RNA polimerase untuk memulai transkripsi.
Untuk memulai transkripsi, faktor transkripsi umum, seperti TFIID, TFIIH, dan lainnya, harus terlebih dahulu berikatan dengan kotak TATA dan merekrut RNA polimerase ke lokasi tersebut. Pengikatan faktor transkripsi pengatur tambahan ke elemen cis- acting akan meningkatkan atau mencegah transkripsi. Selain sekuens promotor, daerah enhancer membantu meningkatkan transkripsi. Enhancer dapat berada di hulu, hilir, di dalam gen itu sendiri, atau pada kromosom lain. Faktor transkripsi berikatan dengan daerah enhancer untuk meningkatkan atau mencegah transkripsi.
Kontrol pasca-transkripsi dapat terjadi pada setiap tahap setelah transkripsi, termasuk penyambungan RNA, pengangkutan nuklir, dan stabilitas RNA. Setelah RNA ditranskripsikan, RNA harus diproses untuk menciptakan RNA matang yang siap diterjemahkan. Proses ini melibatkan penghapusan intron yang tidak mengkode protein. Spliceosom mengikat sinyal yang menandai batas ekson/intron untuk menghilangkan intron dan menggabungkan ekson. Setelah ini terjadi, RNA menjadi matang dan dapat diterjemahkan. RNA dibuat dan disambung di nukleus, tetapi perlu diangkut ke sitoplasma untuk diterjemahkan. RNA diangkut ke sitoplasma melalui kompleks pori nuklir. Setelah RNA berada di sitoplasma, lamanya waktu RNA berada di sana sebelum didegradasi, yang disebut stabilitas RNA, juga dapat diubah untuk mengontrol jumlah total protein yang disintesis. Stabilitas RNA dapat ditingkatkan, yang menyebabkan waktu tinggal lebih lama di sitoplasma, atau diturunkan, yang menyebabkan waktu lebih singkat dan sintesis protein lebih sedikit. Stabilitas RNA dikendalikan oleh protein pengikat RNA (RPB) dan mikroRNA (miRNA). RPB dan miRNA ini berikatan dengan 5′ UTR atau 3′ UTR RNA untuk meningkatkan atau menurunkan stabilitas RNA. Tergantung pada RBP-nya, stabilitas dapat meningkat atau menurun secara signifikan; namun, miRNA selalu menurunkan stabilitas dan mendorong degradasi.
Video pembelajaran: